TÁMOP-4.1.2.E-13/1/KONV-2013-0012
"Sporttudományi képzés fejlesztése
a Dunántúlon"
- -> Tananyagfejlesztés
- -> A vázizom...
- -> 5.


















5.
5. AZ ERŐKIFEJTÉS MECHANIKAI ASPEKTUSAI
- 5.1. Az aktív hossz-feszülés kapcsolat az izomban
- 5.2. A passzív hossz-feszülés kapcsolat az izomban
- 5.3. A az aktív és passzív hossz-feszülés kapcsolat együttes megjelenése
- 5.4. Az ízületi szöghelyzet és az erőkifejtés kapcsolata
- 5.5. Az erő-sebesség kapcsolat az izomban
- 5.6. Az izomrost hosszának és rövidülési sebességének kapcsolata
- 5.7. Elasztikus energia tárolása az izomban
Az eddigi ismereteink alapján beláthatjuk, hogy az izomrostok molekuláris szintű erőkifejtése bonyolult biokémiai és élettani folyamatok eredménye. Vajon mindez hogyan hat hétköznapi, vagy akár sportmozgásunkra, ahol saját testünk ellenállását, vagy külső terheket kell legyőznünk? Mégis hogyan lehetséges az, hogy az ember különböző feladatszituációkban csak kisebb erőt képes kifejteni, vagy esetleg kisebb sebességgel tudja mozgatni végtagjait? Nem szükséges magas szintű mozgástapasztalattal rendelkeznünk ahhoz, hogy megítéljük, milyen mozgásokat, feladatokat tudunk elvégezni, és mindezeket milyen sebességgel. Az idős emberek például tisztában vannak azzal, hogy ha egy alacsonyabb székre ülnek le, akkor onnan nehezebben fognak felállni. Az emberek tapasztalatból tudják, hogy egy teher felemelése akkor végezhető el a legkisebb erőkifejtéssel, ha azt minél közelebb a testközéppontjához teszi meg, csökkentve ezzel a teher felemeléséhez szükséges forgatónyomatékot. A sportolók pedig tudatában vannak annak, hogy nagyobb súlyokat csak kisebb sebességgel tudnak mozgatni. Hogy mindezt megértsük, a következőkben a vázizom erőkifejtésének mértékét befolyásoló mechanikai tényezőket részletezzük.
A teljes izom-ín rendszer kétféle módon tud erőt kifejteni: aktívan és passzívan . Ezeket az erőkifejtési módozatokat talán akkor értjük meg igazán, ha elképzeljük, hogy egy kipreparált izmot két ínvégződésénél rögzítünk. Ha az izmot elektromossággal ingereljük, akkor a kontraktilis fehérjék, vagyis az aktin és miozin közötti kereszthidak létrejönnek, így az ínak a rögzítés helyén húzóerőt fejtenek ki. Amennyiben az izmot nem ingereljük, hanem a két ínnál fogva elkezdjük nyújtani, ellenállást tapasztalunk, tehát az ínak megint csak erőt fejtenek ki a rögzítés helyére. Míg az első szituációban az izom aktívan (active force), ez utóbbiban passzívan (passive force) fejt ki erőt. Az izmok által képződött – akár aktív, akár passzív – húzóerőt a szakirodalom feszülésnek (tension) is nevezi.
5.1. Az aktív hossz-feszülés kapcsolat az izomban
Az izom aktív hossz-feszülés kapcsolatának (length-tension relationship) megértéséhez mindjárt a fentiekben említett példák egyikét használjuk fel. Nemcsak az idős emberek, de bárki elfogadja azt, hogy egy alacsonyabb székről nehezebben lehet felállni, ugyanis valamiért ebben a pozícióban kisebb erőt tudunk kifejteni. Hasonlóan a sportolók is felismerték, hogy bizonyos testpozíciókban jóval kisebb erőt tudnak kifejteni, mint más pozíciókban. A mélyguggolás gyakorlatnál például köztudott, hogy kisebb súlyt tudunk mozgatni, mint a félguggolásnál. Ennek oka az, hogy az ízületek behajlításával és kinyújtásával változik az ízületet áthidaló izom hossza (rövidült, vagy megnyúlt állapotban van), és emiatt változik az erőkifejtő képessége.
Magát a jelenséget az aktív hossz-feszülés görbével jellemzik (5.1. ábra), és a jelenség egyik okát a szarkomerek szerkezetében és működési jellegzetességében találjuk. Korábban már tárgyaltuk, hogy az aktív erőkifejtésért az aktin és miozin filamentumok közötti kapcsolat, a kereszthíd felelős. Huxley „csúszó filamentumok” elmélete szerint kontrakció közben a szarkomerek rövidülnek, mert a párhuzamos aktin és mizoin szálak egymás mellett elcsúsznak a kereszthíd ciklusok révén. Ha egy kipreparált izomrostot annyira szétnyújtunk, amennyire csak lehetséges, akkor viszont a szarkomerek két vége olyan mértékben eltávolodik egymástól, hogy az aktin és miozin filamentumok között megszűnik az átfedés, vagyis nem tud kereszthíd létesülni. Márpedig ha nem létesül kereszthíd, akkor nincs aktív (!) erőkifejtés (aktív hossz-feszülés görbe leszálló ága). Amennyiben a szarkomerek rövidülnek, egyre több kereszthíd tud létesülni, hiszen az aktin és miozin filamentumok átfedettsége egyre nő. Ennek következtében az izomrost erőkifejtése is nő. Amikor a két filamentum teljesen átfedi egymást, akkor maximális számú kereszthíd létesülhet, és az izomrost által kifejtett erő is maximális lesz (aktív hossz-feszülés görbe csúcsa). Ezt a kutatók optimális izomhossznak nevezik. Amennyiben a szarkomerek tovább rövidülnek, a szarkomeren belül olyan kevés lesz a hely, hogy maguk az aktin molekulák is összecsúsznak, egymást átfedve. Az ilyen „dupla átfedésnél” a szarkomer két oldalán elhelyezkedő aktinok zavarják egymást a kereszthídciklusban, így az erőkifejtés ismét csökken (aktív hossz-feszülés görbe felszálló ága). Az izom aktív hossz-feszülés kapcsolata tehát azt jelenti, hogy az izom különböző hossznál eltérően képes aktív erőt kifejteni az aktin és miozin molekulák átfedettségének arányában. Fontos megjegyeznünk, hogy az „aktív” szót azért használjuk, mert itt kizárólag a szarkomer aktin és miozin filamentumainak köszönhető akaratlagos (aktív) erőkifejtést befolyásoló mechanizmust tárgyaltuk. Tudniillik a szarkomerben található passzív (elasztikus) elemek is felelősek az erőkifejtésért, de ezt külön ismertetjük a következőkben.
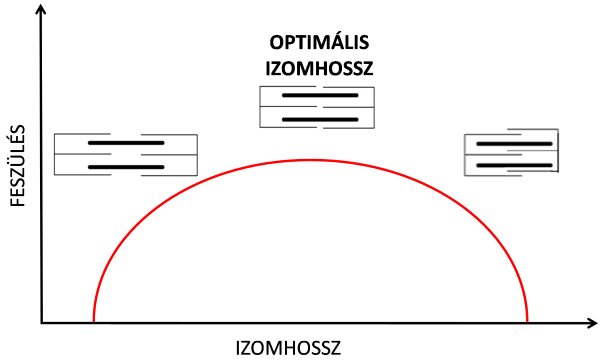
5.1. ábra. Az izom aktív hossz-feszülés görbéje. A piros vonal az aktív erőkifejtés/feszülés mértékét jelöli, mely a szarkomerekben lévő aktin és miozin filamentumok átfedettségének mértékétől függ.
5.2. A passzív hossz-feszülés kapcsolat az izomban
Már korábban említésre került, hogy ha egy izmot passzívan nyújtunk, akkor a nyújtás mértékének arányában fokozott ellenállás tanúsít. Ezt korábban már passzív feszülésnek/erőkifejtésnek neveztük, mely exponenciálisan nő a nyújtás mértékével (5.2. ábra). Az izom passzív feszülése a szarkomerek passzív elasztikus fehérjéinek köszönhető. Bár a titin rendkívül rugalmas fehérje, amely eredeti hosszához képest többszörösére is nyúlhat, a szarkomer szinte valamennyi fehérjére rendelkezik elaszticitással. Az izom tehát a nélkül is képes erőt kifejteni, hogy aktív lenne.
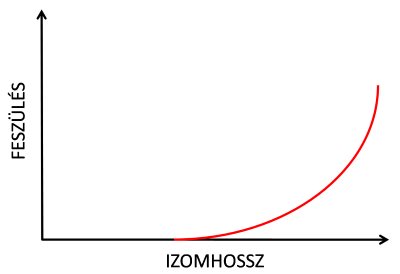
5.2. ábra. Az izom passzív hossz-feszülés görbéje. A passzív feszülésért/erőkifejtésében az izom összes elasztikus fehérjéje, elsősorban a titin játszik szerepet.
5.3. A az aktív és passzív hossz-feszülés kapcsolat együttes megjelenése
Mind az aktív, mind pedig a passzív izomfeszülésnek fontos szerepe van az emberi mozgásokban. Az izmok aktív erőkifejtéssel győzik le a külső és belső ellenállásokat a gravitáció ellen. Ilyenkor az izmok által kifejtett erő nagyságától függ, hogy egy terhet képesek vagyunk-e felemelni, vagy egyáltalán emelt állapotban csak megtartani. Ezzel szemben az izmok passzív erőkifejtése inkább az ízületek stabilizálásában, vagy az aktívan működő izmok lefékezésében, mint antagonista „fék” játszik szerepet. Ha az izmok elveszítenék ezt a passzív rugalmasságot, akkor a mozgások nem lennének annyira kifinomultak.
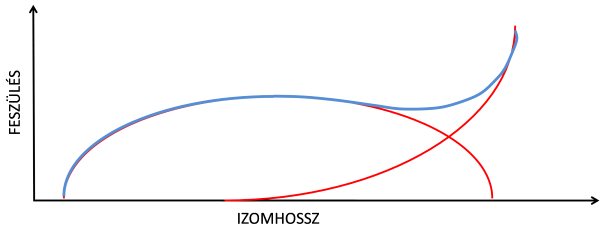
5.3. ábra. A vázizom aktív és passzív hossz-feszülés tulajdonságának együttes hatása. A kék vonal az aktív és a passzív feszülés értékeinek összegét mutatja.
Az aktív és passzív hossz-feszülés görbe kombinációját (5.3. ábra) talán ismét egy kipreparált izom modell alkalmazásával érthetjük meg a legkönnyebben. Ha az izmot egyszerre aktiváljuk is és nyújtjuk is, akkor rövidebb izomhossznál, vagyis a nyújtás elején a feszülést/erőkifejtést csak a kontraktilis izomfehérjék hozzák létre. Ha az aktív izmot tovább nyújtjuk, a szarkomerek annyira szétcsúsznak, hogy az erőkifejtés/feszülés csökken, de még a szarkomer passzív fehérjéi nem nyúltak meg kellőképpen. Még további nyújtás esetén viszont a szarkomer passzív elemei már egyre nagyobb ellenállást tanúsítnak, a feszülés ismét nő.
5.4. Az ízületi szöghelyzet és az erőkifejtés kapcsolata
Bár az izmok eredésüknél és tapadásuknál fogva erőt fejtenek ki, az erőkifejtés következtében az egyes csontokat elmozdítják. A csontok ízesülése mentén forgómozgás jön létre, vagyis az izmok erejüknél fogva egy adott ízületben forgatónyomatékot hoznak létre. A forgatónyomaték (torque) olyan fizikai mennyiség, amely egy erőhatás által egy adott forgáspontban kifejtett forgatóképességet jelenti. A forgatónyomaték a ható erő és az erőkar hosszának szorzatával számolható ki az alábbiak szerint:
M (Nm) = F (N) · k (m)
ahol M = forgatónyomaték, F = erőhatás, k = erőkar hossza. Az erőkar a forgástengelyből az erőhatás útjára merőlegesen húzott szakasz. Az 5.4. ábrán egy kedvező és egy kedvezőtlen ízületi szöghelyzet látható. Bár mindkét esetben az izomerő azonos, a kedvezőtlen szöghelyzetben az erőkar kisebb, így a keletkező M forgatónyomaték is kisebb.
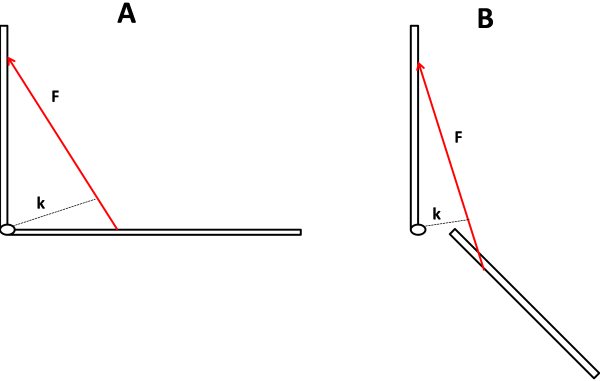
5.4. ábra. Az ízületben keletkező forgatónyomaték kedvező (A) és kedvezőtlen (B) helyzete. F = az izom által kifejtett erő, k = erőkar.
Nem szabad figyelmen kívül hagynunk azt, hogy az ízületi szöghelyzet változásával nemcsak az erőkar nagysága, hanem az izomhossz is változik. És mivel az előzőekben már tisztáztuk, hogy az izom különböző hossz (aktin – miozin átfedettség) esetén különböző erőt fejt ki, ezért egy adott ízületi szöghelyzetben kifejthető forgatónyomatékot nemcsak az erőkar, hanem az aktuális izomhossz is meghatározza. Az emberi szervezetben számtalan ízületi rendszerrel találkozunk és valamennyi sajátos forgatónyomaték – szöghelyzet görbével rendelkezik. Azt az ízületi szöghelyzetet, amelyben a legnagyobb forgatónyomaték érhető el, optimális szöghelyzetnek nevezzük (optimal joint angle). Térdfeszítő izmok esetében ez kb. 60-80 fok, térdflexoroknál pedig 10-20 fok.
5.5. Az erő-sebesség kapcsolat az izomban
Amikor az izom maximális ereje ugyanakkora, mint a legyőzendő ellenállásból adódó erő, akkor az izomban nem jön létre rövidülés, izometriás kontrakcióról beszélünk. Ezt az erőt P0-nak szokás jelölni az erő-sebesség kapcsolat magyarázatakor. Amennyiben csökkentjük az ellenállás nagyságát, az izomerő meghaladja a legyőzendő ellenállás erejét, vagyis az izom rövidül, koncentrikus kontrakciót végez. Megfigyelték, hogy minél kisebb ellenállást kell az izomnak legyőznie, annál nagyobb rövidülési sebességgel képest azt végrehajtani. A felfedezés Archibald Vivian Hill (1938), híres angol fiziológus nevéhez fűződik. Az izom ellenállás nélküli rövidülési sebességét maximális rövidülésis sebességnek nevezzük, jelölése Vmax. Az izom erő-sebesség összefüggése, mely hiperbolikus görbét mutat (5.5. ábra), az alábbiak szerint írhatjuk fel:
(P + a) v = b (P0 – P)
ahol P = aktuális izomerő, P0 = maximálisan elérhető izomerő, v = aktuális rövidülési sebesség. Az a és b kísérletek útján nyert konstansok, melyek az egyes izmokat jellemzik. Értéke általában 0,25. Az egyenlet és a görbe is mutatja, hogy amint növeljük a legyőzendő ellenállás nagyságát, az ellenállás lehetséges mozgatási sebessége nagyságrendekkel csökken. Ugyanez fordítva is igaz: minél nagyobb sebességgel akarunk mozogni, annál kisebb ellenállás esetében tudjuk ezt véghezvinni. Képzeljük el, hogy egy sportoló maximum 100 kg tömegű súlyzót képes félguggolás helyzetben megtartani statikusan. Amennyiben 80 kg-ra csökkentjük a súlyt, képes lesz vele felállni, bár csak lassú sebességgel. Ha még kisebb súllyal terheljük, akkor képes lesz gyorsabban felállni. Egészen kis súllyal (pl. 30-40 kg-mal) már olyan nagy sebességgel képes felállni, hogy akár fel is tud ugrani vele. A Hill-összefüggés egyik legelfogadottabb magyarázata az, hogy nagyobb sebességű kontrakcióknál a kereszthíd ciklusok létrejötte és megszűnése olyan gyorsan következik be, hogy az egyidejű kereszthíd-kapcsolatok száma jelentősen csökken. És kevesebb kerszthíd-kapcsolat kisebb erőkifejtést eredményez. Az erő-sebesség görbe alakja függ az izom architektúrájától és a rosttípus aránytól is. Nagyobb arányú gyors izomrost esetén a görbe laposabb, vagyis az izom képes nagyobb ellenállást is nagy sebességgel mozgatni. A Hill-összefüggés hasznosíthatósága igen jelentős a gyorserőt igénylő sportokban. Ezekben a sportágakban a fő edzéscél a görbe kiegyenesítése, vagyis hogy minél nagyobb ellenállást tudjanak a sportolók gyorsan mozgatni, amely lehet akár a saját testtömeg is (pl. sprinteknél, ugrásoknál). A Hill-görbén azonos ponthoz tartozó erő és sebesség értékek szorzatát mechanikai teljesítménynek (power) nevezzük. A mechanikai teljesítmény kiszámolása tehát az alábbiak szerint történik:
P(W) = F(N) · v(m/s)
ahol P = mechnaikai teljesítmény, F = erő, v = rövidülési sebesség.
A fent említett példánkhoz visszatérve, vajon mi történne, ha a sportoló által legyőzendő ellenállást 100 kg fölé emeljük, vagyis nagyobbra, mint amekkorát izometriásan képes megtartani. 120 kg ellenállást például képes lenne kontrolláltan leengedni, vagyis a sportoló izmai excentrikus kontrakciót végeznének. Az erő-sebesség összefüggés görbéjén látható, hogy amennyiben a maximális izometriásnál még nagyobb erőt kívánunk kifejteni, azt már csak excentrikusan vagyunk képesek, vagyis az izom aktív nyúlásával. Ilyenkor az izomban a kereszthidak nagy része nem válik szét úgy, mint a koncentrikus kontrakcióra jellemző ATP-függő ciklusban, hanem egy darabig ellenállnak a nyújtásnak, és végül a mechanikai erő hatására válnak szét. Ennek köszönhető, hogy általában nagyobb erőt tudunk kifejteni excentrikusan, mint koncentrikusan. Ugyanakkor a mikrosérülések oka is erre a kontrakció típusra vezethető vissza, melynek mechanizmusát a későbbiekben tárgyaljuk. A mai erőfejlesztési módszerek között igen népszerűvé vált az excentrikus erőfejlesztés, ugyanis gyors izomkeresztmetszet- és erőnövekedést lehet vele elérni.
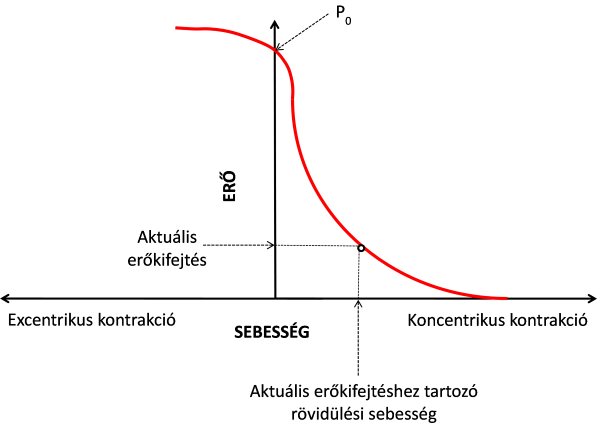
5.5. ábra. Az erő-sebesség összefüggés Hill (1938) alapján. P0 = az izom maximális izometriás ereje.
5.6. Az izomrost hosszának és rövidülési sebességének kapcsolata
Az izomrost anatómiai hossza jelentősen befolyásolja annak maximális rövidülési sebességét. Általában a hosszabb izmok hosszabb izomrostokból állnak, a hosszabb izomrostokban pedig több a sorba kapcsolt szarkomer. Ha elképzeljük, hogy egy izomrostban valamennyi szarkomer egyszerre rövidül, akkor érthető, hogy egy izomroston belül minél több a sorbakapcsolt szarkomer annál nagyobb lesz a rövidülési sebesség. A szarkomerek rövidülése a Z vonalak közeledését és a H-zóna csökkenését jelenti (lásd: csúszó filamentum mechanizmus) és a Z vonalak elmozdulásának nagysága összegződik egy izomrostban (5.6. ábra).
Az orsó alakú izmokban hosszabb rostok találhatók, értelemszerűen ezek gyorsabban képesek rövidülni, mint a tollazott izmok, amelyek jóval rövidebb rostokat tartalmaznak. Általában a flexor izomcsoportok tartalmaznak hosszabb rostokat.
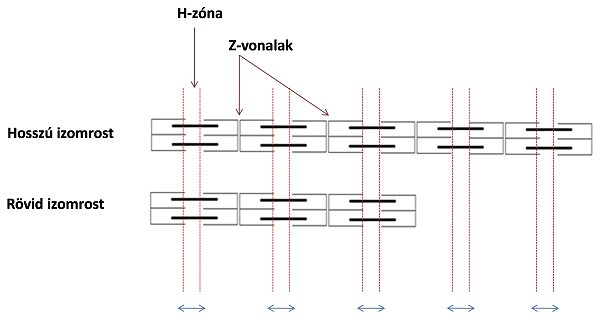
5.6. ábra. A szarkomerek rövidülésének mennyisége (kék nyíl). A hosszabb izomrostokban több szarkomer található, így ugyanannyi idő alatt egyszerre több szarkomer rövidülhet meg: nagyobb a rost rövidülési sebessége.
5.7. Elasztikus energia tárolása az izomban
Megfigyelték, hogy az izom a koncentrikus fázisban nagyobb erő kifejtésére képes akkor, ha azt viszonylag nagy sebességű excentrikus fázis előzi meg (Cavagna és mtsai 1986), ezáltal növelve az izom pozitív munkavégzését. Ennek az egyik oka az, hogy ha az aktív izmot megnyújtják, akkor a koncentrikus fázis elején az izom feszülése nagyobb, mint akkor, ha az izom csak koncentrikusan kontrahálódik. Az izmok passzív, elasztikus elemei nyújtás alatt képesek elasztikus energiát tárolni, amelynek egy részét a koncentrikus fázis alatt vissza lehet nyerni és így az izom nagyobb munkavégzésre lesz képes (Cavagna és mtsai 1986). Mindezek a tényezők lehetővé teszik, hogy edzett egyéneknél az ilyen nyújtásos-rövidüléses ciklust alkalmazva a függőleges felugrás eredményessége előzetes térdhajlítással akár 6 cm-rel is nagyobb lehet (Komi és Bosco 1978), mint statikus guggolás helyzetéből felugorva.
Fontos megemlíteni azonban, hogy a nyújtásos-rövidüléses ciklus létrejöttéhez az excentrikus fázisban az izomnyúlásnak rövid ideig kell tartania ahhoz, hogy az kedvezzen az izom teljesítményének. Ellenkező esetben, amikor az excentrikus fázis viszonylag lassú, akkor a nyújtásos-rövidüléses kontrakció nem jöhet létre, ilyenkor normál excentrikus-koncentrikus kontrakcióról beszélünk. Ez a fajta kontrakció jellemző például egy súly lassú, kontrollált leengedése és felemelése közben.
Hogyan modellezhetnénk a nyújtásos-rövidüléses kontrakciót? Nyújtásos-rövidüléses kontrakció során az izom-ín rendszerre hirtelen akkora erőhatás hárul, hogy ezt az ellenállást csak excentrikus működés során vagyunk képesek „lefékezni”. Ez jól érzékelhető például, ha egy emelvényről leugrunk azzal a szándékkal, hogy a talajfogás után azonnal felugrunk. A nyújtásos-rövidüléses ciklust és az elasztikus energiatárolási képességet a kutatók általában felugrástesztekkel vizsgálják, mint ahogyan azt a fentiekben is láttuk. A nyújtásos-rövidüléses kontrakció modellezéséhez Kopper és mtsai (2014) munkáját használjuk. Az 5.7. ábra illusztrálja az izom-ínrendszer működési modelljét nyújtásos-rövidüléses kontrakció (jelen példában a mélybeugrás felugrás gyakorlat) alatt. A talajfogás pillanatában, de egyes kutatók szerint még néhány milliszekundummal korábban (pre aktiváció; pre-activation) az izom kontraktilis elemei előfeszülnek (kereszthidak létesülnek). A súlypont süllyesztése (excentrikus fázis) úgy megy végbe, hogy a kontraktilis elemekben az aktin-miozin átfedettség nem változik (mondhatnánk, hogy izometriás feszülés létesül), viszont a soros elasztikus elemek jelentősen megnyúlnak. A megnyúlás miatt elasztikus energia tárolódik. A súlypont a legmélyebb pontra történő süllyedése után megkezdi emelkedését (koncentrikus fázis). A koncentrikus fázisban mind az elasztikus, mind pedig a kontraktilis elemekben rövidülés jön létre, amely a felugrást lehetővé teszi. Ráadásul a kontraktilis elemek energiaigényes (ATP függő) koncentrikus munkavégzéséhez hozzájárul az elasztikus elemekben tárolt és felszabaduló elasztikus energia (ami biokémiai energiát nem igénylő folyamat).
A kutatók általában kétféle mélybeugrás tesztet vizsgálnak: a talajfogásnál kicsi, vagy nagy ízületi hajlítással végrehajtott ugrásokat. A kis ízületi hajlításnál a sportoló arra törekszik, hogy a talajfogás után minél rövidebb idő alatt hajtsa végre a felugrást (5.7. ábra). A nagy ízületi hajlítással végrehajtott ugrásnál viszont a sportoló a talajfogáskor jobban süllyeszti súlypontját (5.8. ábra), így a felugrást egy mélyebb pozícióból kezdi meg. Jelen tankönyvben nem foglalkozunk azzal és annak magyarázatával, hogy melyikkel lehet nagyobbat ugrani és miért, mert ez a leugrás magasságától jelentősen függ. Viszont fontosnak tartjuk megjegyezni, hogy a rövid ideig tartó talajkontakt (vagyis az talajfogás utáni azonnali felugrás szándék) során nagyobb az elasztikus energiatárolás (Kopper és mtsai, 2014), mint a hosszabb ideig tartó és nagyobb súlypontsüllyesztéssel végzett ugrásnál. Ez utóbbinál ugyanis kisebb az izom előfeszülése (pre aktivációja), ami miatt a kontraktilis fehérjék kissé eltávolodnak egymástól, és ami miatt kisebb lesz az elasztikus elemek megnyúlása. Számos szerző is javasolja, hogy a pliometriás gyakorlatokat rövid ideig tartó talajkontakttal érdemes végezni az elasztikus energiatárolási képesség növelésére.
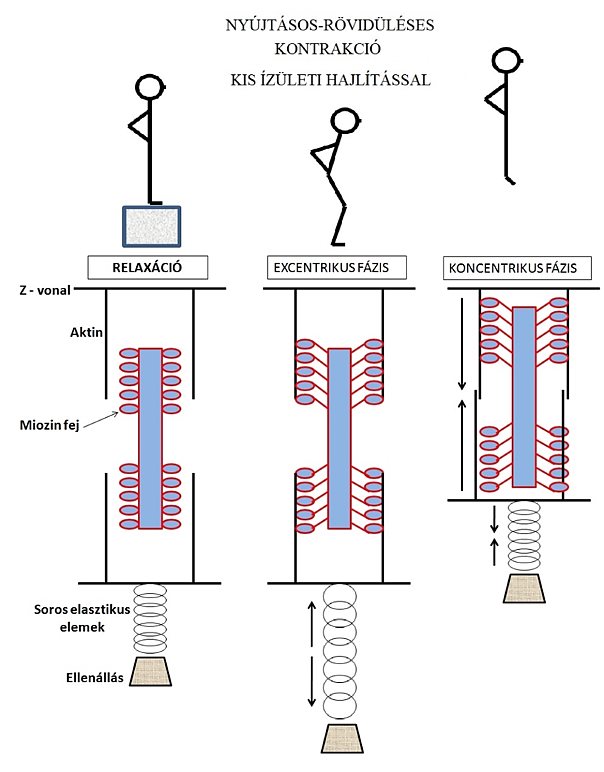
5.7. ábra. Kontraktilis és elasztikus izomrészek modellezése kis ízületi hajlítással végzett mélybeugrás (nyújtásos-rövidüléses kontrakció) közben. Az ábra bal oldalán található izom inaktív, csupán a testtartáshoz elegendő izomerő kerül kifejtésre (a sportoló felkészült a leugráshoz). A középső izommodellben (talajfogás és súlypont süllyesztése) kereszthidak létesülnek, az aktin és miozin filamentumok nem csúsznak el (izometriás előfeszülés), mert jelen ábrán a kis ízületi hajlítással végzett mélybeugrást modellezzük. Az elasztikus elemek megnyúlnak, jelentős elasztikus energiát tárolva. A jobb oldali izommodellben az elasztikus és kontraktilis elemek egyaránt rövidülnek, a súlypont nagy gyorsulással elindul felfelé, megtörténik a felugrás.
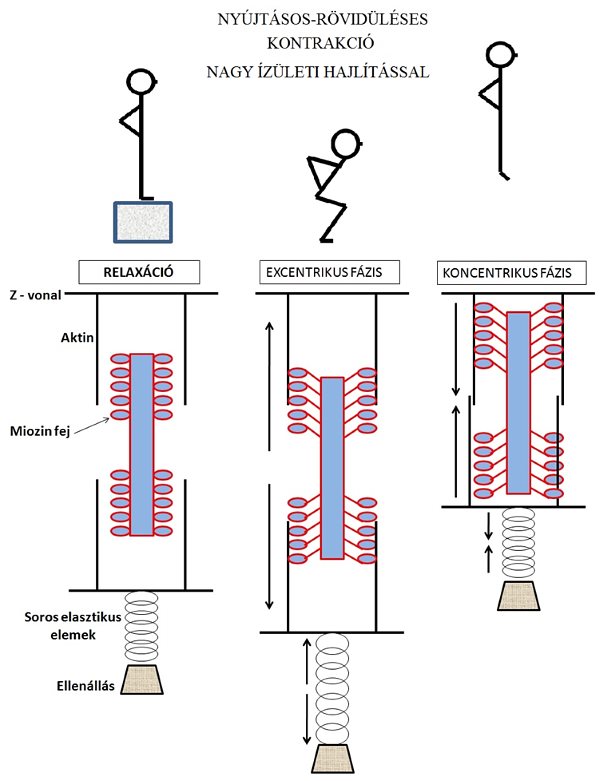
5.8. ábra. Kontraktilis és elasztikus izomrészek modellezése nagy ízületi hajlítással végzett mélybeugrás (nyújtásos-rövidüléses kontrakció) közben. Az ábra bal oldalán található izom inaktív, csupán a testtartáshoz elegendő izomerő kerül kifejtésre (a sportoló felkészült a leugráshoz). A középső izommodellben (talajfogás és súlypont süllyesztése) bár kereszthidak létesülnek, az aktin és miozin filamentumok eltávolodnak egymástól, mert jelen ábrán a nagy ízületi hajlítással végzett mélybeugrást modellezzük. Az elasztikus elemek is kissé megnyúlnak, csekély elasztikus energiát tárolva. A jobb oldali izommodellben az elasztikus és kontraktilis elemek egyaránt rövidülnek, a súlypont nagy gyorsulással elindul felfelé, megtörténik a felugrás.
A fentiekben leírt mechanizmus alapján érzékelhető, hogy a gyors, ciklikus jellegű nyújtásos-rövidüléses kontrakciókból álló mozgások során a nagy mechanikai munkavégzés ellenére csekély az energiafelhasználás. Amikor ugrókötelet hajtunk, a mechanikai munkavégzés nagy, hiszen felugrásokat végzünk, de ehhez képest a mozgás metabolikus igénye kevés (enyhén emelkedik a szívfrekvencia és a tejsavszint). Nagyobb elasztikus energiatárolási lehetőségünk van akkor, ha valamilyen rugalmas felületen végzünk mozgást, például gumiasztalon, vagy ha például kangoo cipővel ugrálunk. Egyes emlősállatok alsó végtagi izomzatához rendkívül hosszú inak párosulnak (pl. impala, antilop), mely a nagy sebességű futások ellenére gazdaságos energiafelhasználást jelent, lehetőséget adva hosszabb ideig tartó menekülésre.
A nyújtásos-rövidüléses ciklust sajátos idegrendszeri tevékenység vezérli. Rövid ideig tartó, de jól időzített elektromos impulzusok lehetővé teszik, hogy az energiafelhasználás minél kisebb legyen (alacsony ATP igény), és az inakban tárolt elasztikus energia minél jobban hasznosuljon. A sportolók által közkedvelt szökdelő- vagy ugrógyakorlatok ezt a képességet kiválóan fejlesztik. Több kutató is igazolta, hogy plyometriás edzés hatására javul a futás gazdaságossága, vagyis egy ilyen program után ugyanakkora távot ugyanakkora sebességgel kisebb oxigénfogyasztás mellett tudunk teljesíteni. Az elasztikus energiatárolási képesség még olyan alacsony sebességű mozgásnál is jelentősen hozzájárul a gazdaságosságához, mint a járás.
ÖSSZEFOGLALÁS
- Az izom aktív erőkifejtése függ az aktuális izomhossztól, mivel eltérő izomhossznál különböző az aktin-miozin átfedés mértéke.
- Az izom a passzív nyújtáskor feszüléssel válaszol, mely a nyújtás nagyságával arányosan nő.
- Minél nagyobb az izom rövidülési sebessége, annál kisebb erőt képes kifejteni. Ezt az izom erő-sebesség összefüggésének nevezzük.
- Mechanikai teljesítmény alatt az izom aktuális rövidülési sebességének és erőkifejtésének szorzatát értjük.
- Hosszabb izomrostok rövidülési sebessége nagyobb, mint a rövideké, mert több sorbakapcsolt szarkomer található benne, melyek egyidejűleg rövidülnek.
Irodalomjegyzék
Cavagna G, Dusman B, Margaria R (1986). Positive work done by a previously stretched muscle. Journal of Applied Physiology, 24, 21-32.
Hill, A.V. (October 1938). "The heat of shortening and dynamics constants of muscles". Proc. R. Soc. Lond. B (London: Royal Society) 126, 136–195.
Komi PV, Bosco C (1978) Utilization of stored elastic energy in leg extensor muscles by men and women. Medicine and Science in Sports and Exercise, 10, 261-265.
Kopper B, Csende Z, Trzaskoma L, Tihanyi J (2014) Stretch-shortening cycle characteristics during vertical jumps carried out with small and large range of motion. Journal of Electromyography and Kinesiology, 24, 233, 239.
| « Előző fejezet | Tartalomjegyzék | Következő fejezet » |
Események
Jelenleg nincs aktuális esemény.