TÁMOP-4.1.2.E-13/1/KONV-2013-0012
"Sporttudományi képzés fejlesztése
a Dunántúlon"
- -> Tananyagfejlesztés
- -> Mozgásszabályozás
- -> 1.


















1.
1. A csontok és izmok felépítése, működése
- 1.1. A csontok felépítése
- 1.2. A csontok alakja
- 1.3. A csontok összeköttetései
- 1.4. A harántcsíkolt izomszövet felépítése
- 1.5. A harántcsíkolt izomszövet működése
- 1.6. Az elektromechanikai kapcsolat, a triád szerkezete és működése
- 1.7. A vázizom kontrakciójának molekuláris mechanizmusa
- 1.8. Az izomműködés mechanikai jellemzői
- 1.9. Energiaforrások az izomműködés során
- 1.10. Hőtermelés az izomban
- 1.11. A vázizomrostok típusai
Mozgás-szervrendszerünk passzív része a csontváz, aktív része a vázizomzat, melyet - tömött rostos kötőszövet - az inak kapcsolnak a csontokhoz.
1.1. A csontok felépítése
Az emberi test stabil belső keretre, a csontvázra épül, melynek alapvető tartóoszlopa a gerinc. A csont a természet alkotta egyik legerősebb anyag, 16,3cm 3 csont kb. 8600kg terhelést képes elviselni, ami legalább öt emelődaru erejével vetekszik. Csontvázunk a teljes testtömeg mindössze 10-20%-át teszi ki, azaz a súlya körülbelül 10kg. Egy hasonló mérettel rendelkező acélrúd ennél ötször nehezebb lenne.
Csontvázrendszerünk alapvető funkciói a következők:
- a test belső szilárd vázát alkotja, meghatározza annak alakját és méreteit,
- elhatárolja és védi a külső behatásoktól a testüregben lévő szerveket,
- szivacsos állományában található a szervezet legfontosabb vérképző szerve, a vörös csontvelő,
- az emberi test mozgásának passzív szerve,
- az emberi szervezet számára fontos ionok, elemek raktára, ahonnan a szükséges anyagok megfelelő folyamatokon keresztül mobilizálhatók, pl. Ca2+.
Az emberi csontvázat a támasztószövetek csoportjába tartozó csontszövet és porcszövet építi fel. A csontok 40%-a víz, a fennmaradó vízmentes rész szervetlen és a szerves alkotórészek adják:
- 35-40% a szerves alkotórész (osszein), melyek a csont rugalmasságát biztosítják
- 60-70% a szervetlen alkotórész, melyek a szilárdságot adják: pl.: Ca2+-foszfát, Ca2+-hidroxiapatit, illetve kisebb részt mészsók, alkálisók, fluoridok, különböző nyomelemek.
Idős korban a szervetlen anyagok részaránya megnő a rugalmasságot okozó szerves alkotók csökkenése miatt, így a kor előrehaladtával rugalmatlanabbá válnak a csontok, törékenységük nagymértékben fokozódik.
1.1.1. A csontok szerkezete
A csontok felépítését egy csöves csonton mutatjuk be (1-1. ábra). A csont külső felszínét rostos burok, a csonthártya borítja. E csontok középső csőszerű darabját, mely tömött csontállományból (kompakt csontállomány) áll, diafízisnek, két vaskosabb végdarabját, epifízisnek nevezzük. A törzshöz közelebb levő végét proximális-, a távolabbit disztális epifíziseknek hívjuk. Az epifíziseket (izületi vég) változó vastagságban üvegporc borítja. Mindkét epifízist szivacsos csontállomány tölti ki, melynek üregeit fiatal korban vörös csontvelő tölti ki, mely az évek múlásával visszahúzódik, helyére sárga csontvelő (zsírszövet) kerül.
A csont középső részének üregét velőüregnek nevezzük, mely felnőtt korban sárga csontvelőt tartalmaz.
*Kiegészítés: A vörös csontvelő zsírszövetté való alakulásával a vörösvérsejt-képző funkcióját elveszti, és csak a szervezetet érő nagyobb trauma (pl.: vérveszteség) hatására aktivizálódik.
1.2. A csontok alakja
Alak szerint csöves, lapos, köbös és labirintusos csontokat különböztetünk meg, melyeket célszerűbb, ha alak szerinti felosztásukban, a magyar kifejezések szerint definiáljuk.
1.2.1. Csöves csontok
Főleg a végtagokban előforduló üreges, cső alakú csontok (szerkezetét lásd előbb). Ilyen, pl.: combcsont, felkarcsont, orsócsont.
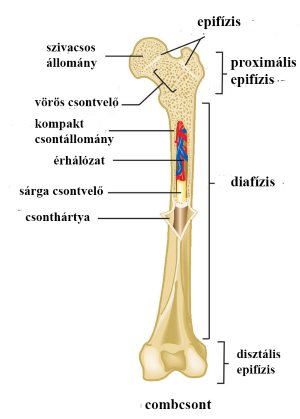
1-1. ábra: A csöves csontok szerkezete
1.2.2. Lapos csontok
Jellemzőjük, hogy az élet során mindig megmaradó vörös csontvelővel teltek, így vérképző szervünk oroszlánrészét adják. Ilyen pl.: agykoponya csontjai, lapockák, csípőcsontok, a bordák, valamint a mellcsont.
1.2.3. Köbös csontok
Szabálytalan, de különböző irányokban alig eltérő méretű csontok. Ilyen pl.: kéz- és lábtőcsontok, ill. a csigolyák testei. Végtagjaink köbös csontjai zsírszövetet, a csigolyatestek vörös csontvelőt tartalmaznak.
1.2.4. Labirintusos vagy légtartalmú (pneumatikus) csontok
Jelentőségük a szerkezet kisfokú könnyítése mellett a hangadás számára fontos rezonálóterek kialakítása. Ilyen pl.: az arcüregben elhelyezkedő homlokcsont.
1.3. A csontok összeköttetései
A csontokat részben rögzített, részben mozgékony összeköttetések fűzik egybe. Abban az esetben, ha a csontok anyaga megszakítás nélkül megy át egymásba, akkor folytonos összeköttetésről, ha az összekötött csontok állománya között nincs meg az állandó anyagfolytonosság, akkor ízületről beszélhetünk.
1.3.1. Folytonos csontösszeköttetések
-
Szalagos csontösszeköttetés
Pl.: lábszár két alsó csontvége, ide sorolhatjuk a koponyacsontok varratos egybeilleszkedését is. -
Porcos csontösszeköttetés
Pl.: csigolya közötti porckorongok, bordák a mellcsonttal. -
Csontos egybeforradás
Pl.: medencecsont a növekedés során, a keresztcsont összecsontosodása öt keresztcsonti csigolyából.
1.3.2. Megszakított csontösszeköttetések
Ízület nek nevezzük azokat az összeköttetéseket, melyeknél a csontfelszínek között rés található. Ízvégeken az egymással ízületi összeköttetésben lévő csontrészeket értjük. Az ízvégek között egyrészt egy domború idomú ún. ízfejet, ill. egy homorút, az ízvápát tudjuk megkülönböztetni, melyek egy zsákszerű kötőszövetbe, az ízületi tokba vannak burkolva. Az ízvégek egymásba illeszkedését szolgáló ízületi felszínek ízületi porccal (üvegporccal) borítottak, melyek megvédik az ízesülő csontvégeket a kopástól.
A csontvégek között néhány cseppnyi nyúlós folyadék, az ízületi nedv (synovia) található, mely jelentősen csökkenti a mozgásokkal járó súrlódást és mintegy kipárnázza az ízületet a rázkódások elviselésére. Az ízesülő csontokat ízületi szalagok rögzítik egymáshoz, melyek szakítási szilárdsága igen nagy. Így pl.: a csípőízület fő szalagja mintegy körülbelül 500kg megterhelésnél szakad el. Laza tokkal és szalagokkal rendelkező, erős izomzattal körülvett ízületekben (pl.: vállízület) az ízvégek egymástól való eltávolodását ficamnak nevezzük, melyet rendszerint az ízületi tok, vagy a szalagok elszakadása kísér.
Az ízületek osztályozhatóak a bennük lévő csontok alapján, így megkülönböztethetünk egyszerű (két csontvég találkozik), ill. összetett (több csont vesz részt) ízületeket.
A/ Egyszerű ízületek
-
Szabad ízületek, melyeket mozgási tengelyeik száma szerint tudjuk csoportosítani
-
Egytengelyű ízületek
- Csuklóízület (hengerízület)
Az elmozdulása a csont hossztengelyére merőleges. Pl.: az ujjpercek között található, szerepe a hajlítás és feszítés. - Forgóízület
Az elmozdulása a csont hossztengelyére egybeesik. Pl.: az 1. és a 2. nyakcsigolya közötti ízület.
- Csuklóízület (hengerízület)
-
Kéttengelyű ízületek
- Tojásízület
Mint a neve is mutatja, tojás alakú ízületi felszínnel rendelkezik, mely két tengely körüli mozgást engedélyez. Pl.: a fej bólogató, ún. „igen-nem” mozgása. - Nyeregízület
Az ízületnek két egymásra merőleges tengelye van. Pl.: hüvelykujj kézközépcsont-kéztőcsont ízülete.
- Tojásízület
-
Soktengelyű ízületek
Elmozdulása három fő- és sok melléktengely mentén történhet. Pl.: vállízület, és a combcsont fejének ízülete.
-
Egytengelyű ízületek
-
Feszes ízületek
Az ízesülő csontvégek kicsit, ill. egyáltalán nem mozoghatnak. Pl.: csípő és keresztcsont közötti ízület.
B/ Összetett ízületek
Olyan bonyolult mozgású ízületek tartoznak ebbe a csoportba, melyek kialakításában kettőnél több csont vesz részt. Pl.: könyök és a térd ízülete.
1.4. A harántcsíkolt izomszövet felépítése
Az emberi szervezetben simaizomszövet, harántcsíkolt izomszövet (vázizom, 1-2. ábra), ill. szívizomszövet található. Mivel a mozgásszabályozás kialakításában a harántcsíkolt izomzat szerepe kimagasló, így a könyvben csak ezen típus felépítésére, működésre térünk ki.

1-2. ábra: A harántcsíkolt izomszövet fénymikroszkópos képe
A mozgás aktív szervrendszerét a vázizomzat adja, mely önálló anatómiai és funkciós egységből, az izmokból áll (1-3. ábra). Az izom húsos részét izomhasnak nevezzük. A vázizmok, melyek a csontváz meghatározott részeiről erednek, és az izomműködés szempontjából fontos, tömött rostos kötőszövetből álló ín segítségével, a csont bizonyos részein tapadnak. Az izmot kívülről kötőszövetes hártya borítja. A hártyából válaszfalak indulnak az izom állományába, melyek az izmot izomnyalábokra tagolják. Az izomnyalábok izomrostokból - azaz sokmagvú izomsejtekből - állnak. Az izomrost a vázizomzat elemi egysége, akciós potenciál generálására képes. Minden egyes izomrosthoz egy speciális szinapszison keresztül (lásd 2.5. fejezet) egy-egy idegrost kapcsolódik. Az idegrost elektromos impulzust juttat tovább az izomrosthoz, melynek hatására az izomrost is ingerületbe kerül (lásd 2.5. fejezet).
Az izomrostokat egy speciális hártya, a szarkolemma borítja. Az izomrostok belsejében ún. miofibrillumok futnak, melyek általában az izomrost hosszával megegyező hosszúságúak. A miofibrillumokat a sejt belső membránrendszere, a szarkoplazmatikus retikulum veszi körül. A miofibrillumok miofilamentumokból épülnek fel, amelyek az elmozduláshoz szükséges fehérjéket tartalmazzák. A vastag filamentumokat miozinnak, a vékonyakat, aktinnak nevezzük. A miozin filamentumnak egy feji- és egy farki részét tudjuk elkülöníteni. A feji részével egyrészt csatlakozni képes a másik miofilamentumhoz, az aktinhoz, másrészt a fej elhajlása okozza a harántcsíkolt izom rövidülését (lásd 1.7. fejezet).
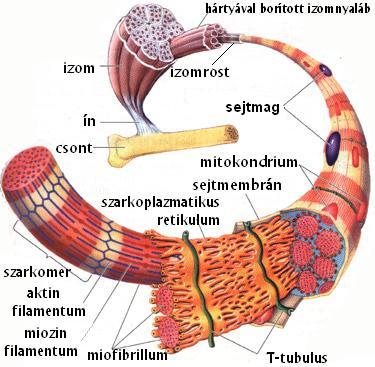
1-3. ábra: A vázizmok felépítése
1.5. A harántcsíkolt izomszövet működése
A harántcsíkolt izmokban az aktin- és miozinfehérjék szabályos rendezettsége alakítja ki a harántcsíkolt jelleget. Megfigyelhető, hogy a rostok hossztengelye mentén sötétebb és világosabb szakaszok váltakoznak. A sötét szakaszt „A” csíknak, míg a világost, „I” csík nak hívjuk. Az „A” csík közepén egy világosabb „H” csík található, közepén az ún. „M” zónával. Az „I” csíkok közepén a „Z” lemezt figyelhetjük meg. A miofibrillum két Z lemez közötti szakaszát szarkomernek nevezzük, melynek hossza nyugalomban 2-2,5µm (1-4. ábra).
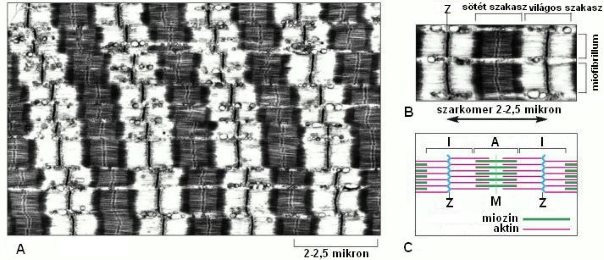
1-4. ábra: A szarkomer elektronmikroszkópos képe (A) és szerkezete (B, C).
A harántcsíkolt izom szerkezetét egyéb stabilizáló fehérjék is alkotják (1-5. ábra). Ezek közül elsőként érdemes megjegyezni a dystrophin nevű fehérjét, mely az aktin és izom-membrán fehérjék közötti kapcsolatot teremti meg. Hiánya a muscularis dystrophia nevű betegséget okozza. Megemlítendők a dezmin, illetve a vimentin fehérjék, melyek az egyes Z lemezeket fűzik össze, valamint a titin, mely a miozin centrális helyzetben tartásáért felelős.
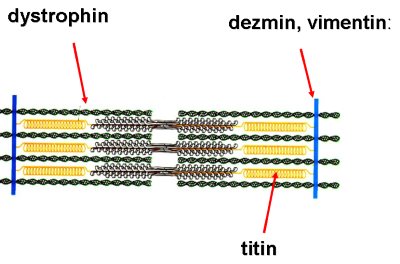
1-5. ábra: A harántcsíkolt izom legfontosabb stabilizáló fehérjéi
1.6. Az elektromechanikai kapcsolat, a triád szerkezete és működése
A vázizomzat működésének alapvető formája az izomrostok kontrakciója. Az izomrostokat körülvevő membránon (szarkolemma) keletkező és terjedő akciós potenciál (lásd később, 2.3. fejezet) és az izom rángása közötti időszakot latenciaidőnek nevezzük, melynek értéke 2-4 ms. Ezen idő alatt azon folyamatok lezajlásához szükséges, amelyek a felszíni membrán potenciálváltozása és a kontrakció kialakulás között mennek végbe. Mindezeket összefoglaló néven elektromechanikai csatolásnak nevezzük. A csatolás első meghatározó lépése az akciós potenciál terjedése az ún. T-tubulusok membránján az izomrost belsejébe.
A T-tubulusok az emlősökben az I- és az A-csík határán elhelyezkedő szarkolemma betüremkedés (1-6. ábra), mely képes akciós potenciál generálására és vezetésére. A vezetési tulajdonságának köszönhetően képes a felszíni membrántól távolabb eső miofribrillumok területére vezeti a potenciálváltozást. A másik rész, mely a szarkoplazmatikus retikulumból áll (L tubulus) kiszélesedett résszel (terminális ciszterna) egészen közel kerülnek a rost belsejébe nyúló T-tubulusokhoz. Emlősökben egy T-tubulushoz két oldalról kapcsolódik egy-egy terminális ciszterna, melyet összefoglaló néven triádnak nevezünk. A szoros kapcsolat ellenére a két struktúra között összeköttetés nem található, membránjaik között mintegy 10-15 nm-es rés mutatható ki. A terminális ciszternák az izomrost Ca2+ raktárának felelnek meg, bennük a Ca2+-ion szabadon, vagy egy speciális fehérjéhez, a kalszekvesztrinhez kötve található. A T-tubulus membránjában egy speciális fehérje, az úgynevezett dihidropiridin (DHP) receptor található. A fehérjemolekula a membránpotenciál változásának hatására szerkezeti változáson megy keresztül, ami elősegíti az L-tubulus terminális ciszternáiban lokalizálódó rianodin (RYR) receptorok megnyílását. Ezek a rianodin-receptorok speciális Ca2+ csatornaként szolgálnak, melyek megnyitása lehetővé teszi, hogy a szarkoplazmatikus retikulumban található Ca2+ raktárakból, a Ca2+ a citoplazmába áramoljon. A kalciumnak a citoplazmába történő kilépése koncentráció-függő folyamat. Nyugalomban a citolpazmában a koncentráció mintegy 100 nmol/l, míg a kontrakció eléréséhez kb. 1 µmol/l koncentrációra történő emelkedés szükséges. A citoplazmába kikerült Ca2+ diffúzióval eljut a Ca2+-kötő helyekhez (lásd 1.7. fejezet), melynek következtében történik meg a harántcsíkolt izom kontrakciója. A citoplazmában a kálciumkoncentráció mindaddig emelkedett marad, míg a Ca 2+ vissza nem kerül a szarkoplazmatikus retikulum területére (nincs ábrázolva). Amennyiben a Ca2+ visszavételét gátoljuk, relaxáció nem tud kialakulni. Ezt a tartós kontrakciós állapotot kontraktúrának hívjuk.
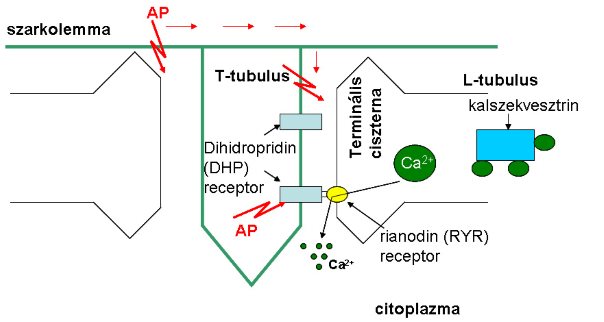
1-6. ábra: A triád szerkezete és a Ca2+-ion felszabadulásának vázlatos rajza
1.7. A vázizom kontrakciójának molekuláris mechanizmusa
Mint ahogy korábban már említettük a vázizomrostok fénymikroszkóppal is jól megfigyelhető szerkezeti sajátossággal rendelkeznek. A vázizomrostok vékony filamentumainak szerkezeti felépítésében legfőképpen az aktin nevű fehérje vesz részt. Az aktin két formája ismert, az egyik a globuláris monomerként azonosított G-aktin, a másik a fibrilláris, több száz monomerből felépülő F-aktin. A vázizmok vékony filamentumait két egymás mellett elhelyezkedő, csavarodott F-aktinlánc alkotja.
A szabályozó fehérjék közül a tropomiozin két aminosavláncból álló vegyület, mely a két aktinlánc között helyezkedik el. A másik szabályozó fehérje a troponin, mely három alegységből áll. A troponin-T alegység alakítja ki a kapcsolatot a tropomiozin fehérjével. A troponin-I alegység, a nevéből is adódóan, szerepet játszik az aktin-miozin kölcsönhatás gátlásában. A harmadik alegység a troponin-C, mely elnevezését onnan kapta, hogy képes a Ca2+ megkötésére. Nyugalomban a tropomiozin-troponin rendszer gátolja az aktinon található miozinkötő helyeket, gátolva ezáltal a kölcsönhatást. Amikor a citoplazmatikus Ca2+ koncentráció a korábban említett módon megemelkedik, a troponin-C alegység kötőhelyei megkötik a Ca2+-ot. A kötődés hatására a komplex konformációváltozáson esik keresztül, melynek következtében szabaddá válik az aktin miozinkötő helyei a vastag filamentumok számára
Az összehúzódáshoz elengedhetetlen az ATP (energiaszállító nukleotid), ill. a korábban említett Ca2+-ionok jelenléte. A kölcsönhatás során a harántcsíkolt izom három állapota figyelhető meg, melyek ciklikus ismétlődése figyelhető meg (1-7. ábra):
- nyugalmi állapot, melynek során a miozinfejhez ATP kötődik, az aktin és miozin nem kapcsolódik egymáshoz,
- aktivált állapot, amikor a miozinfejről az ATP disszociál (leválik) a miozinfej az aktinhoz kötődik (aktomiozin), megtörténik az elmozdulás (ha a miozinfejhez újabb ATP kapcsolódik, a kötés megszűnik, a folyamat előröl indul),
- rigor állapot, akkor alakul ki, ha a szervezetben nincs ATP, ilyenkor a miozinfej nem képes leválni az aktinról, szoros kapcsolat alakul ki (ez az oka a hullamerevség = rigor mortis kialakulásának).
Az aktivált izomban az aktin és miozin között kialakult kereszthidak a vastag filamentummal mintegy 90º-os szöget zárnak be. Az ATP leválásával a kereszthidak kb. 45º-os szöghelyzetbe kerülnek, és közben magukkal húzzák a hozzájuk kapcsolódó vékony filamentumokat („csúszófilamentum mechanizmus – sliding mechanizmus). Mindez azt jelenti, hogy a szarkomert határoló Z-lemezek a szarkomer közepe felé mozdulnak el, változtatva ezzel a szarkomer hosszát. Az aktivált állapot vége és a következő szakasz, a rigor állapot közös jellemzője az átmeneti ATP hiány. Az újabb ATP-molekula kötődése megszünteti az aktomiozin kapcsolatot, és a kereszthidak visszatérhetnek a 90º-os szöghelyzetbe. Amennyiben az aktivált állapot fennáll, azaz a citoplazmatikus Ca2+ szint magas, és a troponin-C alegységek továbbra is Ca2+-ot kötnek, úgy újra létrejöhet az aktin és miozinfej kapcsolódása, a szarkomer rövidülése folytatódhat. Amennyiben időközben az aktiváció megszűnik, az izomrost elernyed, a kereszthidak felveszik a nyugalmi állapotra jellemző szerkezeti elrendezésüket.
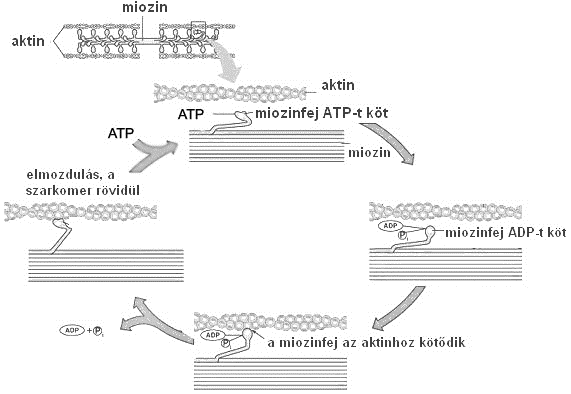
1-7. ábra: A harántcsíkolt izom kontrakciója
1.8. Az izomműködés mechanikai jellemzői
1.8.1. Az izomkontrakció típusai
Egyetlen akciós potenciál során kialakuló mechanikai választ izomrángásnak nevezünk (1-8/a ábra). Az izomrángás időtartama függ az izom típusától. A gyors izomrostokban a kontrakció ideje kb. 7 ms, míg a lassú izomrostokban ez az érték elérheti akár a 100 ms-ot is.
Az izomkontrakció a kontraktilis elemek rövidülésével jár, azonban a kontraktilis elemekkel elasztikus és viszkózus elemek sorba vannak kapcsolva, így a kontrakció úgy is létrejöhet, hogy a teljes izom hossza csökkenne. Az olyan szélsőséges kontrakciót, amikor az izom hossza nem változik, csak a feszülés fokozódik, izometriás kontrakciónak nevezzük (pl nehéz tárgy felemelése). A másik szélsőséges esetben a megrövidülésnek nincs akadálya, a megrövidülés változatlan feszülés mellet következik be. Az ilyen kontrakciót, amikor az izom hossza változik, azonban a feszülés nem, izotóniás kontrakciónak nevezzük. Ebben az esetben akkor, amikor az izom megrövidül anizometriás koncentrikus kontrakció nak, amikor az izom megnyúlik, anizometriás excentrikus kontrakciónak nevezzük. A hossz- és a feszülésváltozás az izmokban leggyakrabban egyidejűleg következik be. Az ilyen kontrakciót ahol az izom hossza és feszülése is változik, auxotóniás kontrakciónak nevezzük.
A megrövidülés sebessége annál nagyobb, minél kisebb erővel szemben kell, hogy végbemenjen. Az izomerő és a megrövidülés sebessége közötti összefüggést az ún. Hill-egyenlet írja le, ami:
(F + a) * (V + b) = (F0 + a) * b = konstans,
ahol F az izomerőt jelenti, V a megrövidülés sebességét, F0 az izom maximális izometriás erejét, a és b az adott izomra jellemző értékek. Mindebből az következik, hogy a feszülés és a megrövidülési sebesség fordítottan arányos, azaz a leggyorsabb hosszváltozás akkor következhet be, ha az izom terhelés nélkül kontrahál, míg a legnagyobb feszülés pedig izometriásan váltható ki. A különböző mozgásokat a sportban ún. dinamikus és statikus gyakorlatokra osztjuk aszerint, hogy az említett izommozgás milyen elemeket (rövidüléses-nyújtásos, izometrikus) tartalmaz.
A vázizomzat esetén nem fordul elő, hogy egyetlen egy akciós potenciál érje az izomrostot, a motoros idegsejteken mindig ingerek sorozata jut el az izomzathoz. Amennyiben az ingerlési frekvencia nő, akkor az izomfeszülés minden egyes rángás után fokozódik, az első kontrakcióra újabb rakódik (1-8/B ábra). Ezt a jelenséget szuperpoziciónak nevezzük. Ennek a lépcsős véltozásnak az oka az, hogy a Ca2+ mennyisége minden egyes ingereknél az előző aktiválásból maradt citoplazmatikus Ca2+ koncentrációhoz adódik, befolyásolva ezzel az aktin-miozin kölcsönhatást. Tovább növelve az ingerlési frekvenciát (15-30/s), az egyes kontrakciók egyre inkább egymásra rakódnak, és a szummálódott ingerek között csak részleges relaxáció tapasztalható, ilyenkor inkomplett tetanuszról (1-8/C ábra) beszélünk. Amikor a sorozatingerlés frekvenciája 30-60/s-ot ér el, az egyes rángások már nem lesznek elkülöníthetőek, elernyedés nélküli, komplett tetanusz (1-8/D ábra) jön létre.
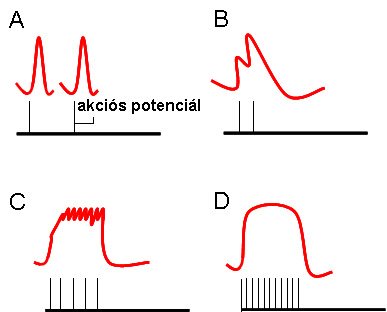
1-8. ábra: A vázizom-kontrakciók formái
A: rángás, B: szuperpozició, C:inkomplett tetanusz, D:komplett tetanusz
1.8.2. Az izomhossz és az izomfeszülés közötti összefüggés
Az izomrost maximális ereje, azaz feszülése az izometriásan kiváltódott komplett tetanusszal jellemezhető. Ha úgy alakítjuk ki a tetanuszos összehúzódásokat, hogy közben változtatjuk az izomrost hosszát, jellegzetes összefüggést kaphatunk (1-9. ábra). A legnagyobb kontrakció kiváltása akkor lehetséges, amikor a rost hossza kb 2 µm-es szarkomerhossznak (optimális szarkomerhossz) felel meg. Amennyiben a szarkomerhossz ennél nagyobb, vagy kisebb, úgy a kontrakciós erő mindkét esetben csökken. A változás oka az, hogy a rostok nyújtásakor a kereszthidak száma változik, ami befolyásolja a generálható erő nagyságát. Igen nagyfokú nyújtás (kb. 4 µm) mellett az átfedés teljesen megszűnik, így ebben az esetben mechanikai válasz nem hozható létre.
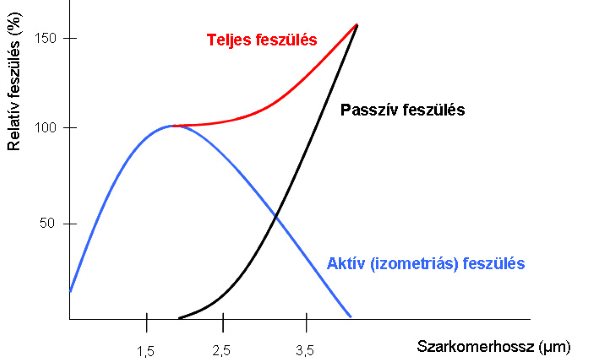
1-9. ábra: A vázizomrost hosszának és feszülésének összefüggése
1.9. Energiaforrások az izomműködés során
Ismert tény, hogy az izomkontrakció energiaigényes folyamat, melynek során az izom a kémia energiát mechanikai energiává alakítja. Az izomműködés közvetlen energiaforrása az energiagazdag szerves foszfátvegyületek képezik, melyek anaerob módon állnak rendelkezésre. A vázizom a kémiai energia kb. 40-50%-át tudja mechanikai energiává alakítani, a többi hőtermelés formájában valósul meg. Az így felszabadult hő jelentősen hozzájárul a szervezet hőszabályozásához. Az aerob és anaerob folyamatok arányai fontosak és egyik sem kizárólagos, hiszen ismert, hogy még álmunkban is termelünk tejsavat, ami az életfolyamathoz szükséges.
1.9.1. Anaerob energiaképzés
Az izomműködés közvetlen energiaforrása az adenozin-trifoszfát (ATP), mely nyugalmi körülmények között csak mmol/l-es koncentrációban található a citoplazmában, így csak időlegesen tudja fedezni a kontrakcióhoz szükséges energiát. A raktározott ATP, mindösszesen néhány mp-ig képes fedezni az energiaszükségletet, azonban az ATP folyamatos reszintézise fenntartja a szükséges energiaellátást. A reszintézis forrásaként említendő akreatin-foszfát (kreatin-P, ugyancsak néhány mmol/l). A kreatin-foszfát nem közvetlenül felhasználható energiaforrás, azonban egy enzim, a kreatin-foszfokináz képes a kreatin-P + ADP (adenozin-difoszfát) ↔ kreatin + ATP reakció alatt az időközben kimerült ATP tartalék pótlására. A kreatin-foszfát kb. 5-6 másodpercnyi energiabiztosítást jelent, így a foszfátvegyületek mindösszesen 7-10 mp munka energiaigényét képesek fedezni anaerob alaktacid módon. Nagy intenzitású sprintteljesítménynél (100 m-es síkfutás) előfordul, hogy a teljes munkát anaerob módon végezzük. Néhány másodperces munkavégzés után a foszfátraktárak teljesen kiürülnek. Az ennél hosszabb munkavégzésnél kerül sor a szénhidrátok, jelen esetben a glikogén anaerob módon történő felhasználására (anaerob laktacid). A folyamat első lépése a glikogenolízis, mely folyamatban a glikogén glükózzá alakul. Ezután a glükóz a citoplazmában, a glikolízis alatt több lépcsőn keresztül, enzimek segítségével, három szénatomos piroszőlősavvá alakul. Amennyiben az oxigénellátás nem kielégítő, a keletkezett piroszőlősav nem lép be a mitokondriumba és tejsavvá (C3H6O3) redukálódik. A keletkezett tejsavat, illetve annak nátrummal vagy káliummal létrejött sóját (laktát) a vérkeringés a májhoz juttatja, ahol a glikogén reszintézise újra megtörténik. Ezt a folyamatot Cori-körnek hívjuk. Ilyen módon egy molekula glükóz bontásából 2 ATP molekula képződik, mely jelentősen elmarad az aerob módon történő energianyeréstől.
1.9.2. Aerob energiaképzés
Az aerob energianyerés forrásai a szénhidrátok, a zsírok, és a fehérjék lehetnek. A szénhidrátok a már korábban említett módon piroszőlősavvá alakulnak. A keletkezett vegyület oxigén jelenlétében acetilcsoporttá alakul, koenzim-A (KoA) molekulához kapcsolódik (AcKoA), és a mitokondriumba szállítódik. A két szénatomos acetilcsoport a citrátciklusba (Szentgyörgyi-Krebs ciklus) belépve az oxálecetsav vegyülethez kapcsolódik. A körfolyamat végtermékeként szén-dioxid (ezt lélegezzük ki) és nikotinamid-adenin-dinukleotid (NADH) keletkezik. A NADH hidrogénionjából (H+) a terminális oxidáció ciklus alatt az oxigénhez kapcsolódva víz keletkezik. Mindebből adódik, hogy egy molekula glükóz égése során további 36 mol ATP képződik, így összesen, a glikolízist beleértve 38 mol, melynek egyenlete a következő:
C6H12O6 + 6O2 =6CO2 + 6H2O + 38 ATP
A szervezet a legnagyobb energiaraktárát a zsírok biztosítják. A zsírok kizárólagosan csak aerob módon tudnak energiát szolgáltatni, anaerob módon a zsírokból nem lehet energiát nyerni az izomműködéshez. A zsírok nagy része, három szénatomos, úgynevezett trigliceridek formájában raktározódnak a szervezetben. A b-oxidáció folyamatában a trigliceridekből 2 szénatomos egységek, acetilcsoportok keletkeznek. Ezek a csoportok a KoA molekulához kapcsolódnak, aminek segítségével a mitokondriumhoz jutnak, majd a citrátciklusba lépnek. Mint ahogy korábban említettük, a mitokondriumban lezajló citrátciklus megfelelő oxigénellátottság mellett képes ellátni a feladatát, így ezek a folyamatok csak aerob körülmények között képesek végbemenni. Energetikai szempontból a zsírok bontása a szénhidrátokból származó energia többszörösét jelenti, ugyanis amennyiben egy trigliceridhez 3 db 18 szénatomos zsírsav (sztearinsav) kapcsolódik, abban az esetben a triglicerid lebontásából mintegy 330 mol ATP keletkezik. Mivel a zsírraktáraink gyorsan pótlódni képesek, így a szervezet energiahiánya csak rendkívüli esetben áll fenn, mint néhány extrém aerobkapacitást igénylő sportágaknál, mint pl. országúti kerékpárosoknál, maratoni futóknál. Mindezek miatt a szénhidrátszintjüket ezeknél a sportolóknál folyamatosan szinten kell tartani a fizikai aktivitás során.
A fehérjék a zsírokkal megegyezően kizárólagosan csak aerob módon képesek bomlani. Egészségesen táplált szervezetben a fehérjék bontására nem kerül sor, mert először a szénhidrátok és zsírtartalékainkat használjuk fel. A fehérjék aminosavakból épülnek fel. A bontásukkor először az aminocsoport lehasítása történik meg (deaminálás), majd a szervezet a maradék szénláncot hasznosítja. A lehasított aminocsoportok karbamidként (urea) a vizelettel távoznak a szervezetből.
1.10. Hőtermelés az izomban
Az izomműködés során a kémiai energia egy része hőenergiává alakul. A keletkezett hőenergiának különböző részeit lehet megkülönböztetni. A nyugalmi hő, nyugalomban lévő izmon mérhető, mely az alapanyagcsere-folyamatok során keletkezik. A kezdeti hő a kontrakció alatt termelt hőmennyiség, mely az aktiválási hőből, illetve a megrövidülési hőből tevődik össze. Az aktiválási hő az izom kontrakciójakor szabadul fel, a megrövidülési hő nagysága arányos az izomrövidülés hosszával. A kezdeti hő aerob és anaerob körülmények esetén egyaránt megfigyelhető, ami elsősorban a kreatin-foszfát bontásával magyarázható. A kontrakciót követően a hőtermelés akár 30 percig is folytatódhat, amit regenerációs hőnek is nevezünk. A regenerációs hő nagysága nagyjából megegyezik a kezdeti hővel, azaz a regeneráció alatti hőtermelés megegyezik a kontrakció alatt termelt mennyiséggel.
A harántcsíkolt izom hőtermelési folyamata esszenciálisnak mondható, hiszen az izomműködés az egyetlen olyan folyamat, amivel a szervezet jelentősen növelni tudja a hőtermelését. Minden esetben izommunkát kell végeznünk, ha a szervezetet kihűlés veszélyezteti.
1.11. A vázizomrostok típusai
Az emberi harántcsíkolt izmok rostösszetétele kevert. Körülbelül 200 éve ismert, hogy vannak sötét, illetve világos rostok. A tudomány fejlődése révén a harántcsíkolt izmok felosztása megváltozott (1-1. táblázat) melyek a következők lehetnek:
Azok a rostok, melyek lassú, tónusos összehúzódásra képesek, nem fáradékonyak, és kitartó munkát képesek végezni ST (slow twitch), I. típusú, vagy más néven lassú oxidatív (slow oxidatív, SO) rostoknak nevezzük. Főbb jellemzőik, hogy sok mioglobint sok mitochondriumot tartalmaznak, gazdag a kapillárishálózatuk. Színük alapján vörös rostoknak is nevezzük. Nagy számban találhatók a testtartást biztosító nyak- és törzsizomzatban.
Az izomrostok másik típusát azok a rostok (FT, fast twitch), II. típusú alkotják, melyek gyors, fázisos, robbanékony, azonban hamar fáradó összehúzódásra képesek. Ebben a csoportban két alcsoportot lehet elkülöníteni: (i) az ún. II.B típusú rostokat, melyek gyors glikolízissel (FG, fast glycolitic: gyors glikolitikus) energiát nyerő rostok, alacsony mioglobin tartalmúak, kevés mitochondriumot, kevés kapillárist és sok glikogént tartalmaznak. Színük fehér, ATP-t anaerob úton állítanak elő, nem tudják folyamatosan ellátni az izmot elegendő ATP-vel, könnyen kifáradnak, az ATP-t gyorsan bontják és gyors kontrakciós sebességűek, azonban nagyon fáradékonyak. A másik alcsoportba (ii) az ún.II.A típusú rostokat soroljuk, melyek oxidatív és glikolitikus energiaendszerrel is működő ( fast oxidative glycolitic: gyors, oxidatív glikolitikus, FOG), sok mioglobint tartalmaznak, sok bennük a mitochondrium és gazdag a kapillárishálózat. Színük ’piros’, magas kapacitással szintetizálnak oxidatív úton ATP-t, az ATP-t gyorsan bontják, közepes erőkifejtésre képesek, és közepes gyorsasággal fáradnak.
| I. típusú izomrost (SO) | II.A típusú izomrost (FOG) | II.B típusú izomrost (FG) | |
| Szín | vörös | ’piros’ | fehér |
| Mitokondrium | nagyon sok | sok | kevés |
| Rostátmérő | vékony | közepes | vastag |
| Z lemez | vastag | közepes | vékony |
| Kapillárisok | nagyon sok | sok | kevés |
| Glikolitikus aktivitás | kevés | közepes | nagy |
| ATP-áz aktivitás | kevés | közepes | nagy |
| Mioglobintartalom | sok | közepes | kevés |
| Kontrakció sebessége | lassú | gyors | gyors |
| Fáradékonyság | lassan fárad | közepese fárad | gyorsan fárad |
1-1. táblázat: Izomrosttípusok és legfontosabb jellemzőik
Az edzéstípus helyes megválasztásával az izomrostok képesek tanulni, módosulni. A Ila rostok pl. aerob edzés hatására megtanulhatnak oxigént felhasználni, így részben át tudják vállalni a lassú rostok aerob munkavégzését. Ismert, hogy a sprintereknél az II.B rostok az edzések utáni pihenő időszakban felszaporodnak a combizomzatban. Állóképességi edzés esetén a II.B rostok II.A típusúvá, a II.A pedig I. típusú rostokká alakulnak, erőedzés hatására a II.B rostok II.A típusúak lesznek. Nagy intenzitású erőedzés és rövid intervallumú gyors munkavégzés kombinációjával az I. rostok II.A típusúvá válása érhető el (1-10. ábra).
A rostösszetétel meghatározására izombiopszia után elvégzett hisztokémiai vizsgálatok adhatnak választ. Az invazív (véres, szöveti károsodással járó) módon nyert szövetmintát lefagyasztják, majd vékony metszeteket készítve, és megfestve mikroszkóp alatt tanulmányozzák. Az izombiopszia segítségével megállapítható, hogyan változik az izomrostok összetétele gyors fizikai terhelés, vagy különböző edzésformák hatására.
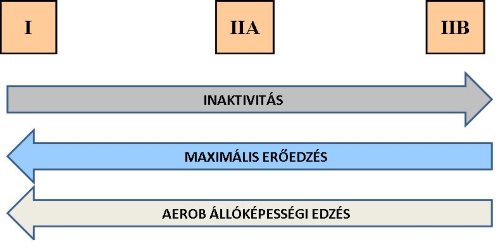
1-10. ábra: Különböző rostátalakulási tendenciák eltérő edzéstípusok következtében.
I: I típusú rost, IIA: II/A típusú rost, IIB: II/B típusú rost.
| « Előző fejezet | Tartalomjegyzék | Következő fejezet » |
Események
Jelenleg nincs aktuális esemény.