TÁMOP-4.1.2.E-13/1/KONV-2013-0012
"Sporttudományi képzés fejlesztése
a Dunántúlon"
- -> Tananyagfejlesztés
- -> Mozgásszabályozás
- -> 3.


















3.
3. Az idegrendszer szerveződési szintjei
- 3.1. A gerincvelő anatómiája
- 3.2. Gerincvelői pályák
- 3.3. Gerincvelő idegek (nervi spinales)
- 3.4. Gerincvelői vázizomreflexek
- 3.5. Az agytörzs
- 3.6. A kisagy
- 3.7. A köztiagy
- 3.8. A nagyagy
- 3.9. Az agyidegek
- 3.10. A limbikus rendszer
- 3.11. Vegetatív idegrendszer
A gerincesek idegrendszerét anatómiailag és működésileg különböző csoportokra oszthatjuk.
Anatómiai felosztás szerint:
- központi idegrendszer, mely a gerincvelőből és agyvelőből (agytörzs, kisagy, köztiagy, nagyagy) épül fel,
- perifériás idegrendszer, ami a gerincvelői idegekből (31 pár) és agyidegekből (12 pár) áll.
A gerincvelői idegek alapvető felépítését és működését ld. a gerincvelő anatómiája (3.1.) és a gerincvelői vázizomreflexek című (3.3.) fejezetekben. Az agyidegek anatómiájával a 3.9. fejezetben foglalkozunk.
Működési felosztás:
- szomatikus idegrendszer, mely magába foglalja az idegrendszer érzőműködéseit, és akaratlagos vagy automatikus mozgatóműködéseit (lásd 4. fejezet),
- vegetatív idegrendszer, többnyire nem tudatosuló, a vegetatív szervek működését (pl.: keringés, légzés, emésztés) szabályozó idegrendszer (lásd 3.11. fejezet).
3.1. A gerincvelő anatómiája
A gerincvelő vagy gerincagy (medulla spinalis) kisujjnyi vastagságú, a gerinccsatornában elhelyezkedő üreges szerv (3-1. ábra). A testből jövő információk nagy része a központi idegrendszer e szakaszán halad át és feldolgozásuk már ezen a szinten megkezdődik. A gerincvelő felszínét az agyhártyához hasonló kötőszövetes burkok borítják, melyek a csontos koponyától való védelmet biztosítják. Emlősökben három burkot tudunk elkülöníteni, melyek belülről-kifelé haladva a következők:
- lágy agyhártya (pia mater), ami közvetlenül a gerincvelő felszínét borítja, két lemezből áll, melyek között öblök (szinuszok) találhatóak,
- pókhálóhártya (arachnoidea), mely hozzáfekszik a kemény agyhártyához,
- kemény agyhártya (dura mater), egy erős rostos lemez, ami a gerinccsatorna belső felszínét borítja.
A lágy-, ill. a pókhálóhártya között keskeny hasadék található, melyet szubarachnoideális térnek is nevezünk. Ezt a teret az agy-gerincvelői folyadék (liquor cerebrospinalis) tölt ki.
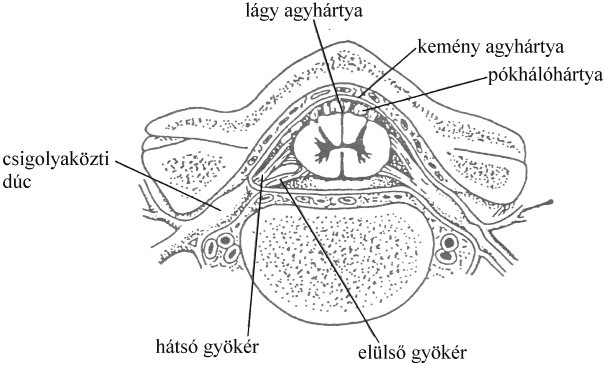
3-1. ábra: A gerincvelő elhelyezkedése a gerinccsatornában
/Forrás: Csoknya, Az idegrendszer összehasonlító morfológiája, 1998./
A gerincvelő kezdete az öreglyuknál található és az első (férfiaknál) vagy második (nőknél) ágyékcsigolya magasságában, fonalszerű maradványban végződik. A gerincvelőt, a gerincoszlophoz hasonlóan különböző szakaszokra oszthatjuk (3-2. ábra). Ennek alapjánnyaki (cervicalis), háti (thoracalis), ágyéki (lumbalis), keresztcsonti (sacralis) és farki (coccygealis) régiókról beszélhetünk. A különböző régiókban a szelvények száma fajonként eltérő. Emberben 8 nyaki, 12 háti, 5 ágyéki, 5 keresztcsonti, 1 csökevényes farki szelvényt találhatunk.
*Kiegészítés: Csak érdekességként említjük meg, hogy az emlősökön belül, a gerincvelő szelvényezettsége nem mutat számottevő eltéréseket, azaz pl. az emberre jellemző 8 nyaki szelvény más emlősállatban pl. a zsiráfban is ugyanúgy megtalálható.
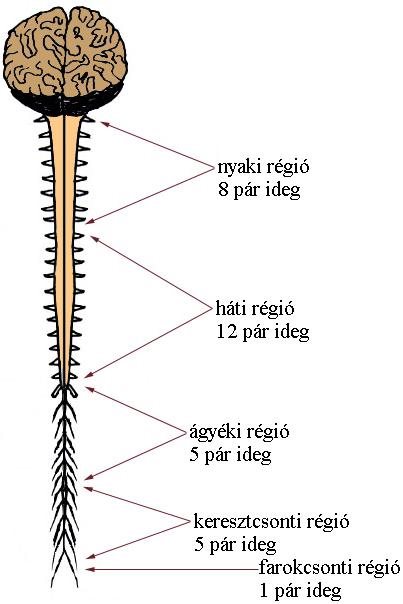
3-2. ábra: A gerincvelő szakaszai
A gerincvelő keresztmetszetén (3-3. ábra) két részt tudunk elkülöníteni, a belső szürkeállományt (substantia grisea), és az azt körülvevő fehérállományt (substantia alba). Elülső felszínén egy mély hasadék (fissura mediana anterior ), míg a hátsó felszínén egy kis barázda található. A szürkeállomány pillangóhoz („H” betű) hasonlítható, melynek közepén aközponti csatorna (canalis centralis) található. A szürkeállomány hátsó szára a gerincvelő hátsó szarva ( cornu posterius), míg az előrefelé kiugró, valamivel vaskosabb rész az elülső vagy mellső szarv (cornu anterius). Az elülső hasadék és a szürkeállomány között az ún. fehér összekötő ereszték (comissura alba) található. A gerincvelő háti és ágyéki szakaszán a hátsó és elülső szarv között oldalt is található egy kiemelkedés, amit oldalsó szarvnak (cornu laterale) nevezünk, melynek a vegetatív idegrendszerben van szerepe. A szarvak térben oszlopokat (columna) alkotnak.
A gerincvelői idegek hátsó-, ill. mellső gyökéren keresztül a gerincvelőből erednek, majd minden gerinces állatban egyesülve, kevert gerincvelői idegeket (érző-, ill. mozgatórostokat egyaránt tartalmaz) alkotnak. A hátsó gyökéren, a hátsó szarvba belépő érzőneuronok sejtteste a csigolyaközti dúcban (ganglion spinale) található. A hátsó szarv mindig érző-, az elülső szarv mindig mozgatóneuronokat tartalmaz. Közöttük lehetnek a szürke állományt el nem hagyó interneuronok (3-4. ábra).
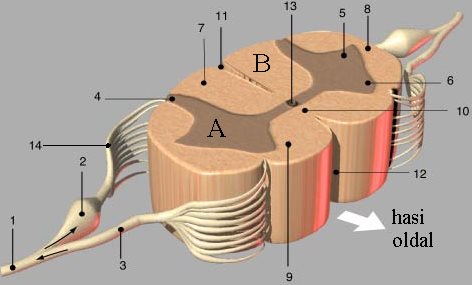
3-3. ábra: A gerincvelő metszetének térbeli modellje
A: szürkeállomány, B: fehérállomány; 1, gerincvelői ideg; 2, csigolyaközti dúc;
3, elülső gyökér; 4, hátsó szarv; 5, oldalsó szarv; 6, elülső szarv;
7, hátsó köteg;8, oldalsó köteg; 9, elülső köteg; 10, fehér összekötő ereszték;
11, hátsó barázda; 12, elülső hasadék; 13, központi csatorna; 14, hátsó gyökér.
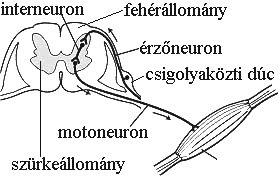
3-4. ábra: Az érzőneuronok, az interneuronok és a motoneuronok elhelyezkedése a gerincvelőben
A szürkeállományt rétegekre, úgynevezett laminákra lehet osztani. A felosztásnál azt kell figyelembe venni, hogy a neuronok sejttestjei hol helyezkednek el. Ennek alapján 10 laminát tudunk elkülöníteni, melyeket római számokkal jelölünk. A rétegek számozása a hátsó szarvtól kezdődik az elülső szarv felé.
A szürkeállomány neuronjait a szerint is csoportosíthatjuk, hogy az axonjaik merre hagyják el a szürkeállományt. Ennek alapján elkülöníthetünkradiculáris vagy gyökérsejteket, melyek axonjai az elülső gyökéren keresztül hagyják el a gerincvelőt. Megkülönböztetünkfuniculáris vagy kötegi neuronokat, melyek axonjai az oldalsó kötegbe lépnek. Végezetül comissurális vagy eresztéki neuronokat ismerünk, melyek axonja a fehér eresztékben kereszteződnek a gerincvelő területén.
A szürkeállományt körülölelő fehérállomány felépítését a következő fejezetben tárgyaljuk.
3.2. Gerincvelői pályák
3.2.1. A fehérállomány szerkezete
A gerincvelő fő feladata az összeköttetés biztosítása a központi idegrendszer alacsonyabb és magasabb szintjei között. Ez alapján a kívül elhelyezkedő fehérállomány (substantia alba) nagy részét hosszában futó, velőhüvelyes rostok képezik. A fehérállományt - a gerincvelő mindkét felében - az onnan ki-, és belépő rostok kötegekre (funiculus) osztják (3-3. ábra), így megkülönböztetünk:
- elülső köteget (funiculus anterior), ahol az egyik legfontosabb felszálló pálya a tractus spinothalamicus található. A másik fontos pálya, ami itt található, a keresztezetlen piramispálya (tractus corticospinalis directus), ami egy leszálló mozgatópálya.
- oldalsó köteget (funiculus lateralis), ahol fel- és leszállópályákat egyaránt találunk. Itt láthatjuk a kisagyba futótractus spinocerebellaris dorsalist és ventralist. Ezen a területen lokalizálható a keresztezett piramispálya ( tractus corticospinalis cruciatus), melyek rostjai a gerincvelő mellső szarvi motoneuronoknak adják át az ingerületet. Az oldalkötegben találhatóak a leszálló vegetatív pályák is, melyek a magasabb központokból induló motoros pályákat tartalmazzák, melyek az oldalsó szarvi vegetatív sejtekhez futnak.
- hátulsó köteget (funiculus posterior), melyek pályái az epikritikus szenzibilitásért (mély nyomás, vibráció, helyzetérzékelés) felelős információkat tartalmazzák. Mediálisan a Goll-féle pálya, laterálisan a Burdach-köteg található
A különböző kötegeken belül, számos rostnyaláb-csoportot tudunk elkülöníteni, melyek a központi idegrendszer gerincvelő pályáit alkotják. A gerincvelő pályarendszereit asszociációs, felszálló (afferens) és leszálló (efferens) csoportra tudjuk felosztani:
3.2.2. Az asszociációs pályák
A gerincvelőből eredő és a gerincvelőben végződő rövid pályák tartoznak ebbe a csoportba.
3.2.3. Felszálló (afferens) pályák
Ezek a pályák a gerincvelőt kötik össze az agy különböző régióival:
- Goll-Burdach, mely a bőrből, izmokból, ízületekből származó érzőingerületeket szállít a nyúltvelőig (Goll-Burdach magok, lásd 3.5. fejezet), ahol végződik, ill. átkapcsolódik, s a talamuszon keresztül a nagyagykéregbe jut,
- Flechsig-féle nyaláb, ami az izmoktól, inaktól közvetít ingerületet a kisagyhoz,
- Gowers-féle nyaláb, az ín-, illetve izomorsókból továbbít ingerületet a kisagyba,
- Edinger-féle nyaláb, ami a legfontosabb érzőingerületet szállító pálya. Fájdalom- és hőérzeteket szállít a talamuszon keresztül a nagyagykéregbe.
3.2.4. Leszálló (efferens) pályák
Ezek a pályák az agy különböző szakaszait kötik össze a gerincvelő motoros sejtjeivel:
- piramispálya (tractus corticospinalis), mely a nagyagykéreg frontális lebenyének piramissejtjeiből indul ki (lásd 4.1. fejezet),
- agytörzsi sejtcsoportokból kiinduló másnéven extrapiramidális pályák (tractus vestibulospinalis, reticulospinalis, olivospinalis, tectospinalis, rubrospinalis) (lásd 6.1. fejezet),
- fájdalomérzést szabályozó leszálló pálya.
3.3. Gerincvelő idegek (nervi spinales)
A gerincvelő idegek a környéki idegrendszer részét képezik. Számuk 31 pár, ebből 8 nyaki, 12 háti, 5 ágyéki, 5, kereszttáji, 1 farki idegpár. Egy-egy gerincvelői ideghez tartozó részt szegmentumnak nevezünk. Az egyes gerincvelői szegmentumok által beidegzett bőrterületetdermatómának nevezzük. A dermatómák érzéskieséséből tudunk következtetni a gerincvelői szegmentumi sérülések lokalizációjára. Egy gerincvelő szelvényen azt értjük, ahonnan egy pár gerincvelő ideg ered. A gerincvelő idegekre az alábbi jellemzők érvényesülnek:
- páros idegek, azaz a gerincvelő szelvényből a jobb és baloldalon 1-1 ideg lép ki, melyek egy dorzális (ramus dorsalis) és egy ventrális ( ramus ventralis) ágra oszlanak,
- kevert idegek, melyek érző és mozgató rostokat egyaránt tartalmaznak,
- két gyökérrel erednek, a hátsó gyökér a hátsó szarv, míg az mellső gyökér az elülső szarv végén ered. A hátsó gyökérben érzőrostok futnak, itt található a csigolyaközti dúc, a mellső gyökérben mozgatórostokat találunk. A mellső gyökér gerincvelő háti- és ágyéki szakasz idegeiben vegetatív szimpatikus rostok is haladnak. Ezek a vegetatív rostok két összekötő ággal (fehér ág, ramus communicans albus, illetve szürke ág, ramus communicans griseus) ún. vegetatív határkötegi dúcokhoz kapcsolódnak,
- szimmetrikus lefutásúak, a test középvonalát nem lépik át.
3.4. Gerincvelői vázizomreflexek
A gerincvelői reflexek lehetővé teszik, hogy bizonyos egyszerűbb mozgási funkciók automatikusan végbemenjenek, agykérgi irányítás és tudatosulás nélkül. A reflex nem más, mint az adott stimulusra automatikusan bekövetkező ingerválasz.
Az idegrendszer működési egysége a reflexív, mely az ingert felfogó receptorokból, az afferens szárból, a központból, az efferens szárból, valamint a végrehajtó szervből, az effektorból áll. A gerincvelői reflexek íve a gerincvelőben záródik, de ez nem jelenti azt, hogy a reflexet kiváltó inger nem kerül észlelésre magasabb szinten (agykéreg), illetve, hogy a legmagasabb szintű központok nem hatnak vissza le a reflexív működésére. Beállítják és szabályozzák izmaink tónusát, feszítettségét, szabályozzák testtartásunkat, és mozdulataink kivitelezését. Általában csak nagyon pontosan meghatározott inger válthat ki egy adott reflexet. Az adott reflex kiváltásához szükséges legkisebb ingert adekvát ingernek nevezzük. Adekvát inger pl. a kutya számára a rovar mászása, mely vakarózási reflexet indít be. Ugyanakkor, ha a többszörös ingerek nem esnek egybe, akkor a vakarózás elmarad. A bolhák ugrálva közlekednek, így az ugrások elválasztják egymástól az érintési ingereket, és nem keletkezik adekvát inger a vakarózáshoz, a reflex nem alakul ki.
A következőkben a legfontosabb reflexeket ismertetjük:
3.4.1. Nyújtási (izomeredetű, saját, vagy miotatikus) reflex
A reflex elsősorban az antigravitációs izmokban jelentkezik. A nyújtási reflex receptora az izomban elhelyezkedő izomorsó, mely az izom megnyúlását, feszülését érzékeli.
Az izomorsók, melyek 4-10 mm-es hosszúságú, kötőszövetes tokkal rendelkező képletek, az izomra ható feszítésről tájékoztatják a központi idegrendszert. Bennük kétfajta afferens idegvégződés található, melyek a feszítettség dinamikus és statikus összetevőinek analízisét végzi. Az izomorsók belsejében intrafuzális izomrostok helyezkednek el, melyek két végükön tartalmaznak kontraktilis részeket. A középső nem kontraktilis szakasz kizárólag csak a nyújtásra érzékeny szenzoros végződéseket tartalmazza. Az izomorsók belsejében háromféle intrafuzális rost található: (i) dinamikus magzsákrost (ii) statikus magzsákrost (iii) néhány magláncrost. Az afferens axonok (Ia, II típusú) végződései spirálisan hálózzák be az intrafuzális rostok középső részét. Az izomorsók a tényleges izomkontrakciót létrehozó extrafuzális rostokkal párhuzamosan helyezkednek el. Amikor az extrafuzális izmok összehúzódnak az izomorsók feszítettsége csökken. Kisegítő mechanizmus hiányában az izom kontrakciója alatt az izomorsók nem érzékelnék az izom feszítettségét, melyre a motorikát koordináló idegrendszeri struktúráknak alapvető szükségük van. Az izomorsók érzékenységét egy speciális mechanizmus, az ún. g-efferensek (Ag-efferens rostok) segítik, melyek az intrafuzális rostok összehúzódását hozzák létre, ezzel szabályozzák az izomorsók nyújtással szembeni érzékenységét. Akaratlagos izomműködés esetén egyidőben aktiválódnak a motoros működésért felelős Aa-, illetve az izomorsó érzékenységét beállító Ag-neuronok.
Az izomorsókból az érzőinformációk a csigolyaközti dúcokban elhelyezkedő ganglionsejtek közreműködésével a gerincvelő szürkeállományába jutnak. Az érzőneuronok axonjai közvetlenül Aa-motoneuronokkal szinaptizálnak a szürkeállomány elülső szarvában. A motoneurok végül visszaküldik axonjaikat abba az izomba, ahonnan a nyújtási inger érkezett és összehúzzák azt. Ez a legrövidebb reflexkör az emberi szervezetben, rövidebb köre mindössze két idegsejtből és az őket összekötő szinapszisból áll (monopszinaptikus reflex). Az érzőneuron axonjának másik ága egy interneuron közbeiktatásával az ellentétesen működő (antagonista) izmot relaxálja (3-5. ábra/A). A miotatikus reflex szerepe az antigravitációs mozgások kivitelezése és a mozgáskoordináció. Ide tartozik pl. a közismert és az orvosi diagnosztikában gyakran használt patellareflex. A térdkalács alatti ínra (ligamentum patellae) reflexkalapáccsal ütést mérve a comb feszítőizma megnyúlik, majd az reflexkör lefutása után ugyanezen feszítő izmok összehúzódnak és az alszár „kirúg”. A nyújtási reflex antigravitációs feladatait úgy látja el, hogy fenntartja a test egyensúlyi helyzetét ülő, vagy álló testhelyzetben, vagy akár mozgás közben is. Képzeljük el, hogy egy álló ember teste kissé előrebillen. Ilyenkor a láb hátsó felszínén lévő izmok megnyúlnak (pl. a háromfejű lábikraizom), majd a miotatikus reflex hatására gyorsan összehúzódnak, és ezáltal hátrafelé döntve visszahozzák a testet a kiindulási egyensúlyi testhelyzetbe.
3.4.2. Inverz nyújtási reflex
Ez a reflex az előzővel éppen ellentétes működésű. A túlnyújtást és az ebből fakadó sérülést hivatott kiküszöbölni (tehermentesítő reflex). A reflex az izom túlzott megnyúlására, elernyedéssel válaszol. Receptora a Golgi-féle ínorsó, mely az izom és az ín találkozásánál, az ín kollagén rostjai között helyezkedik el, és az izom megnyúlását érzékeli. Az inorsók - ellentétben az izomorsókkal – sorosan kapcsolódnak az extrafuzális izomrostokhoz. Az extrafuzális izomrostok megrövidülése meghúzza az inorsóban található kollagénrostokat, és az ott található axonokban (Ib afferens) akciós potenciál keletkezik.
A központba érkező érző rost gátló interneuronon (glicinerg neuron) keresztül csökkenti a motoneuron aktivitását, így az izom, amelyből az ingerület kiindult, relaxálni fog. Az antagonista izom összehúzódik. Erős inger esetén, az ingerület képes átkereszteződni a gerincvelő másik oldalára is (reflex irradiáció), ahol a feszítő izmok összehúzódnak (3-5. ábra/B).
3.4.3. Flexor reflex
A flexor reflex az előzőekkel ellentétben a testre kívülről érkező információkat dolgozza fel, receptorai a bőrben elhelyezkedőszabad idegvégződések, melyek a fájdalmat érzékelik. A fájdalominger a gerincvelőbe érkezve, többszörösen átkapcsol ( poliszinaptikus reflex). Több interneuronon keresztül gátolja az azonos oldali feszítő izmokat, míg serkenti a hajlítókat, így a végtag a veszélyes ingertől távolító mozgást végez. Az ellenkező oldali végtagizmokat ellentétesen aktiválja (keresztezett extenzorreflex), úgy, hogy a feszítőizmokat összehúzza, míg a hajlítókat relaxálja, ezáltal megakadályozza a test egyensúlyának elvesztését (3-5. ábra/C).
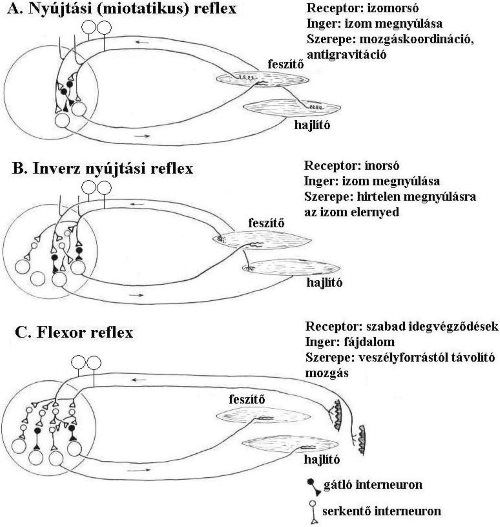
3-5. ábra: Gerincvelői vázizomreflexek sematikus ábrája
(a szürke- és fehérállomány nincs külön ábrázolva)
A: nyújtási (miotatikus) reflex; B: inverz nyújtási reflex; C: flexor reflex
3.5. Az agytörzs
Az agytörzs a gerincvelő folytatása. Részei a nyúltvelő (nyúltagy), a híd és a középagy (3-6. ábra). A z agytörzs hátsó felületén egy mélyedés található, melyet rombus ároknak (fossa rhomboidea) nevezünk. Ez az árok a IV. agykamra.
3.5.1. A nyúltagy szerkezeti egységei és funkciói
- A nyúltagyi piramisban kereszteződnek át a piramispálya axonjai,
- itt található a Goll- és a Burdach-mag (nucleus gracilis és nicleus cuneatus), illetve a lemniscus medialis melyek a gerincvelői felszállópályák fontos állomásai. Ez a pályarendszer szállítja az epikritikus szenzibilitásért (finom tapintás, végtagok helyzete, mozgása) felelős információkat. A pálya a talamusz nucleus ventralis posteromedialis (VPM) és nucleus ventralis posterolateralis (VPL) magjain átkapcsol, majd a szomatoszenzoros kéreg (gyrus postcentralis) Brodmann 3,1,2 areáin végződik,
- a mellső és oldalsó köteg határán található az olajkamag (oliva inferior), mely az extrapiramidális rendszer egyik fontos központja. Innen indul ki a gerincvelő felé a tractus olivospinalis pálya, illetve a kisagy felé a korábban említett kúszórost, a tractus olivocerebellaris,
- itt hagyja el az agyat a IX., X., XI. és a XII. agyideg (lásd 3.9. fejezet),
- itt található a keringésszabályozás két központja a vérnyomásfokozó (presszor) és a vérnyomáscsökkentő (depresszor) központ,
- a légzésszabályozás két fontosabb központja is itt helyezkedik el, a belégzést megindító belégzési ritmus generátor és a gátló vágusz sejtek, melyek leállítják a belégzést, így megindulhat a kilégzés,
- a nyúltvelő-híd határán találhatóak a vesztibuláris magok, ill. a hídba is átnyúló formatio reticularis (agytörzsi hálózatos állomány), melyek a helyzetérzékelésben és az izomtónus kialakításában fontosak (lásd 6.1. fejezet). A formatio reticularis részének tekinthetők a keringési - és légzésközpontok, ill. a nyelési - és hányási reflexek központjai is. Az innen kiinduló pályarendszer (tractus reticulospinalis) az extrapiramidális vezérlésben játszik szerepet.
3.5.2. A híd szerkezeti egységei és funkciói
- A hídkarok összekötik a kisagyat a nagyaggyal és a kisagyat a gerincvelővel (3-6. ábra),
- magasabb rendű légzésközpontok találhatók itt, melyek képesek a nyúltvelői légzésközpontok működését agykérgi tudatos parancsra felülbírálni, mely fontos lehet, pl. a beszédhangok létrehozásakor,
*Kiegészítés: Keringési központok nincsenek a hídban, ezért a légzéssel ellentétben, vérnyomásunkat kevésbé tudjuk akaratlagosan (agykérgi parancsra) befolyásolni.
- itt lép ki az V., VI., VII. és a VIII. agyideg (lásd 3.9. fejezet),
- a formatio reticularis a nyúltvelőből kiindulva egészen a hídig húzódik, szerepeit lásd a nyúltvelőnél korábban,
- itt található az agy szerotonin gyártó központja a raphe-mag, innen a gerincvelőbe leszálló pályák indulnak, melyek részt vesznek a fájdalomérzések csillapításában, ill. felszálló pályák is indulnak az agykéregbe és a hippokampuszba, melyek a memória, a motivációk és sok más magasabb rendű agyi működés szabályozásában vesznek részt.
3.5.3. A középagy szerkezeti egységei és funkciói
- A középagy felső része a tectum, mely főleg egyszerű látóinformációkat (fényerőérzékelés, mozgásérzékelés) feldolgozó agyterület. Az innen kiinduló pálya (tractus tectospinalis) az extrapiramidális rendszer egy része,
- itt található a középagyi vörösmag (nucleus ruber) melyből fontos gerincvelői leszállópálya (tractus rubrospinalis) indul ki. Szerepe van a testtartás kialakításának szabályozásában (lásd 6 1. fejezet),
- fontos része a középagyi feketemag (substantia nigra), mely a bazális ganglionok közé tartozik. A feketemag két szövettani részre a pars compactara és a pars reticulatara osztható. A pars compacta idegsejtjeinek egy része dopamint termel, melyek egy speciális pályán keresztül jutnak el a striatumba, ahol befolyásolják annak működését. Mindezek alapján a pars compactanak az extrapiramidális mozgásszabályozásban van szerepe (lásd 4.2. fejezet), valamint az itt található dopamin tartalmú idegsejtek sérülése okozza a Parkinson-kórt. A pars reticulata a törzsdúcrendszer kimeneti magja a talamusz felé. Funkciói közé tartozik az automatikus mozgások szabályozása, a szemmozgások koordinálása, az orientáció, illetve a tekintet stabilizációja. Parkinson-kór esetében ezek a területek is sérülnek.
3.6. A kisagy
A kisagy (cerebellum) a legfontosabb mozgáskoordináló agyterület. Szerteágazó funkciói miatt több témában is utalni fogunk rá, valamint ebben a fejezetben röviden összefoglaljuk felépítését és működését.
3.6.1. A kisagy anatómiája
A kisagy a hídnál a kisagykarokkal kapcsolódik az agytörzshöz (3-6. ábra). Két féltekéje van, melyeket középen a féreg (vermis) választ el egymástól. A két félteke külső részén tekervényezett szürkeállomány figyelhető meg. A belső fehérállományban találhatók akisagyi mély magvak. A kisagy alapjához csatlakozik a kisagy ősibb evolúciós eredetű része a flocculus (pelyhecske) és a nodulus (csomócska).
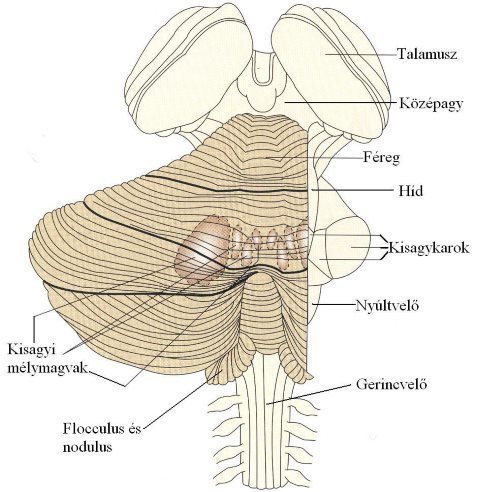
3-6. ábra: A kisagy, mögötte az agytörzs, illetve a talamusz elhelyezkedése hátulnézetben, a két nagyagyfélteke eltávolítása után
3.6.2. A kisagy sejtszintű működése
A kisagy funkcionális egysége a kisagyi modul (3-7. ábra). A kisagyba érkező információk moharostokat és kúszórostokat alkotnak, melyek közül a moharostok interneuronokon (granuláris sejt, kosársejt) keresztül kapcsolódnak a kisagykéreg legfőbb sejtjeihez, a Purkinje sejtekhez, míg a kúszórostok közvetlenül érik el azokat. A kisagyba futó rostok a következő területekről hoznak információkat:
- helyzetérzékeléssel kapcsolatos információk átkapcsolás nélkül, közvetlenül a vesztibuláris idegeken (tractus vestibulocerebellaris),
- szintén helyzetérzékelési információk a nucleus vestibularis inferiorból,
- felszálló érzőinformációk a gerincvelőből az izmok tónusáról (tractus spinocerebellaris),
- információk a nagyagykéreg összes lebenyéből (cortico-ponto-cerebellaris),
- információk a formatio reticularis-ból.
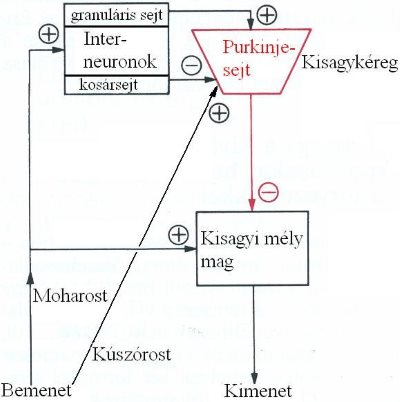
3-7. ábra: A kisagyi információáramlás vázlata
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 864.oldal, 35-24. ábra alapján/
A granuláris sejtek serkentik, míg a kosársejtek gátolják a Purkinje-sejtek működését. A Purkinje-sejtek axonjukkal ezután lejutnak a kisagyi mélymagokhoz és gátolják azokat. A kisagyi mélymagok axonjai adják a kisagy kimeneteit, melyek elsődlegesen a formatio reticularis-ba, a középagyon keresztül a talamuszba, ill. a középagyi vörösmagba jutnak.
*Kiegészítés: Gyakorlatilag kisagyi mélymagnak tekinthető a kisagyon kívül-, a nyúltvelő-híd határán elhelyezkedő Deiters-mag (nucleus vestibularis lateralis) is, mivel közvetlen bemenetet kap a flocculus és a nodulus Purkinje-sejtjeiből. A Deiters-mag az egyik legfontosabb testtartást kialakító központ (lásd 6.1. fejezet).
3.6.3. A kisagy főbb funkcióinak csoportosítása
A kisagy funkcióit három csoportba oszthatjuk, ez a három feladat szorosan kötődik három jól elkülönülő kisagyterülethez (3-8. ábra).
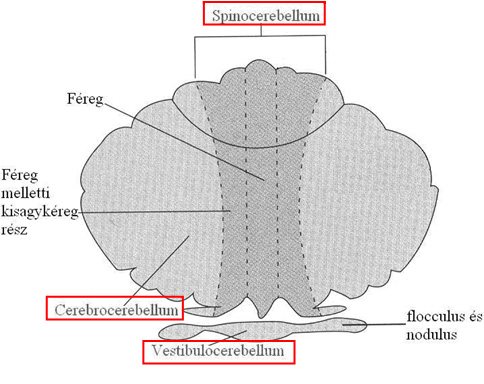
3-8. ábra: A kisagy funkcionálisan elkülönülő részei (piros kerettel jelölve)
A/ Vestibulocerebellum:
A kisagy alapjánál elhelyezkedő flocculus és nodulus alkotja a vestibulocerebellumot, mely nevéből adódóan az egyensúlyérzékelésben, a helyzetérzékelésben, és a testtartás szabályozásában játszik szerepet. Közvetlen kapcsolatban áll a Deiters-maggal, információkat kap a vesztibuláris érzékszervből és a nucleus vestibularis inferiorból.
B/ Spinocerebellum:
A spinocerebellumot a féreg és a közvetlenül mellette lévő kisagykéreg rész alkotja. Szerepe a mozgáskoordináció: a tervezett mozgásokat hasonlítja össze a már kivitelezettekkel, és szükség esetén korrekciós utasításokat közvetít az izmok felé. Bemeneteket kap az agykéregből (mozgástervek) és felszálló információkat a gerincvelőből (izmok állapota).
C/ Cerebrocerebellum
A kisagyféltekék oldalsó része alkotja a cerebrocerebellumot. A kisagy legfejlettebb része, a legtöbb információt képes tárolni. Ennek megfelelően feladata a mozgások tanulása és a már megtanult mozgások rögzülése. A tanult bonyolultabb mozdulatsorok indítása is innen történik, pl. az írás folyamata során, az extrapiramidális pályarendszer segítségével (lásd 4.2. fejezet). Az akaratlagos mozgások tervezésében jelentős premotor agykéreggel és a középagyi vörösmaggal van közvetlen kapcsolatban.
3.7. A köztiagy
A köztiagy a középagyat a két nagyagyféltekével összekötő, a III. agykamra két oldalán elhelyezkedő agyterület. Fontosabb részei az epitalamusz, a talamusz és a hipotalamusz.
3.7.1. Az epitalamusz
Legfontosabb része a tobozmirigy, mely fényérzékelő bemeneteket kap, és sötétben szerotonint alakít át melatoninná. A napsütéses órák csökkenésével párhuzamosan nagy mennyiségben felhalmozódó melatonin depressziót okozhat. Ezt a tünetegyüttest skandináv depressziónak nevezzük.
3.7.2. A talamusz
Ha az agyat egy számítógéphez hasonlítanánk, akkor a talamusz lenne benne a processzor (’tudat kapuja’). Feladatai rendkívül sokrétűek. Általánosan elmondhatjuk, hogy az érzékszervekből érkező információkat szelektálja és sok a gondolkodással, figyelemmel kapcsolatos műveletet végez. Minden érzékszervből érkező információnak - kivétel a szaglás - itt van az utolsó agykéreg előtti átkapcsoló állomása. A talamusz dönti el, mely információk fontosak a gondolkodáshoz, és melyek lényegtelenek. A nem fontos információkat ezután „törli”, a fontosakat pedig ún. reverberációs körökre helyezi (lásd 6.5. fejezet), melyek alkotják a rövidtávú memóriát és belőlük később, hosszú távú emlékek képződhetnek.
*Kiegészítés: A szaglóinformációk a talamusz érintése nélkül is elérhetik az agykérget. A talamusz szerepe így kiesik, ezért tapasztalhatjuk azt, hogy szagokra kevésbé szelektál az agy. Míg pl. egy nem fontos hangingerre nem figyelünk oda, addig minden szaginger eljut a tudatunkig és befolyásolja viselkedésünket. Nem véletlen, hogy a testünk által kibocsátott illatanyagok, a feromonok, döntően meghatározhatják egymás iránti negatív vagy pozitív érzelmeinket, ill. szexuális magatartásunkat.
3.7.3. A hipotalamusz
A hipotalamusz a talamusz alatt helyezkedik el. Az agy legfontosabb vegetatív szabályozó központja. Itt találhatók többek között a szervezet hőháztartását szabályozó hűtő és fűtőközpontok, valamint az éhség- és jóllakottság központja. Nagysejtes magvai az oxitocin és a vazopresszin nevű hormonokat termelik, míg kissejtes magvai az agyalapi mirigy hormontermelését szabályozzák. A hipotalamusz az agyalapi mirigy segítségével eljuttathatja az agy limbikus rendszeréből (lásd 3.8. fejezet) érkező információkat a hormontermelő mirigyekhez. Így közvetítésével valósul meg az idegrendszer és a hormonrendszer kapcsolata. A hipotalamusz segítségével lehetséges az, hogy érzelmi állapotunk változásai - a hormonrendszer közvetítésével - kihathatnak belső szerveink működéseire.
3.8. A nagyagy
A nagyagy az emberi agy legnagyobb tömegű része. Két féltekéből áll melyeket a kérgestest (corpus callosum) köt össze. A féltekéknek van egy domború (3-9. ábra), egy belső (3-10. ábra) és egy alapi felszíne. Külső része a szürkeállomány, tartalmazza az idegsejtek sejttesteit, míg alatta helyezkedik el a fehérállomány, mely az idegsejtek velőhüvelyes axonjait foglalja magába. A fehérállományon belül találhatók még elkülönült szürkeállományú magcsoportok. Ezeket bazális ganglionoknak nevezzük, szerepük főleg az extrapiramidális mozgásvezérlésben van (lásd 4.2. fejezet). A nagyagy felszínén tekervényeket (gyrusok) ésbarázdákat (sulcusok) találhatunk, melyeknek egy része minden egyénben egyforma elhelyezkedésű, ezek négy fontosabb lebenyre tagolják az agyfelszínt (3-9. ábra).
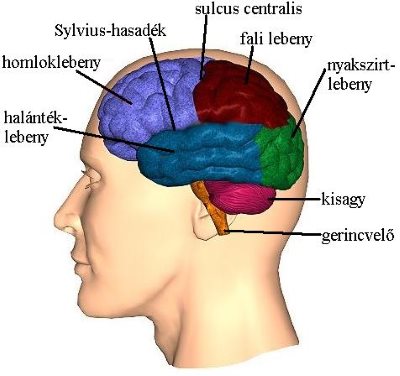
3-9. ábra: A nagyagykéreg lebenyei
3.8.1. A nagyagy lebenyei
Homloklebeny (lobus frontalis)
A fali lebenytől egy árok, a sulcus centralis, választja el. Leghátsó régiója a primer motoros kéreg (gyrus praecentralis), mely az akaratlagos mozgásvezérlés legfőbb központja (lásd 4.1. fejezet). E terület része a bal agyféltekében a Broca-área, mely a beszéd és az írás mozgató központja. Elülső régiójában található az agy egyik legfontosabb asszociációs működésű régiója a prefrontális kéreg (PFC), mely az érzetek kialakulásában, a tanulásban és a személyiség kialakításában fontos (lásd még a limbikus rendszernél, 3.10. fejezet).
Fali lebeny (lobus parietalis)
A sulcus centralistól hátrafelé helyezkedik el. Elülső része az elsődleges érzőkéreg (gyrus postcentralis), mely a bőreredetű érzékelés központja.
Halántéklebeny (lobus temporalis)
A halántékcsont mögött a Sylvius-hasadék alatt helyezkedik el. Többek között a hallóinformációk feldolgozását végzi, ill. általában e területek mellett található a bal féltekében a beszédértő központ, vagy más néven a Wernicke-área. Emellett a halántéklebeny része az egyik legősibb agykéregterület a hippokampusz (3-10. ábra), mely a hosszú távú memória kialakulásában vesz részt (lásd 6.4. fejezet), és egyben a leggyakoribb epilepszia gócpont.
Nyakszirtlebeny (lobus occipitalis)
A nyakszirtlebeny a nagyagy hátsó része, mely a látóinformációk feldolgozását végzi.
3.8.2. Az agykéreg felépítése
Az agykérget 52 areára (Brodmann 1-52) lehet felosztani, melyek a rétegek számában, belső szerkezetükben különböznek. Megkülönböztetjük az ősi kéregtípust, az úgynevezett paleocortexet. Ilyen például a hippokampusz. A másik típushoz a neocortex tartozik, mely hat rétegből épül fel, melyek a következők:
- molekuláris réteg (stratum moleculare): idegrostokat és szinapszisokat tartalmaz,
- külső szemcsés réteg (stratum granulosum externum): kisméretű idegsejtek találhatóak itt,
- kis piramissejtek réteg (stratum pyramidale externum),
- belső szemcsés réteg (stratum granulosum internum): kisméretű idegsejtek találhatóak itt,
- nagy piramissejtek réteg (stratum pyramidale internum),
- plexiform réteg (stratum multiforme): változó alakú és méretű sejtek.
Az agykéreg működési egységei oszlopokba (modul) rendeződnek. Egy-egy modul kb. 200-300 µm átmérőjű, 2.5-3 mm magas, mely magába foglalja az agykéreg valamennyi rétegét. Egy modulhoz nagyjából 5000 sejt tartozik. A humán nagyagy általában 2 millió modult tartalmaz (patkány: 1000, majmok: 1 millió), illetve egy modul kb. 100-200 másikkal tart kapcsolatot. Minden modulba kétféle afferens érkezik. A specifikus afferensek az érzékszervek felől, de már kellőképpen átdolgozott információt szállítanak. A nem specifikus afferensek más kérgi modulokból (corticocorticalis afferensek) a már feldolgozott ingerületet hozzák. A kimenetek (efferens rostok) vagy projekciós rostok lehetnek, melyek elhagyják az agykérget, vagy asszociációs rostok, melyek más modulokhoz szállítanak ingerületet.
Az érzékszervek felől az érzőpályákon érkező rostok, az ún. specifikus afferensek szigorúan a IV. rétegben ágazódnak el. Az itt található sejtek axonja a felületes rétegben szinaptizál a piramissejtek csúcsi dendritjével. A piramissejtek axonjai a kimenetet képviselik. A más modulokból érkező corticocorticalis afferens a kérgi henger tengelyében halad és ágazódik el, közvetlenül végződik a piramissejteken. Az információfeldolgozás a látszólagosnál bonyolultabb a gátló és serkentő interneuronoknak köszönhetően.
3.9. Az agyidegek
Az agyidegeket érző-, mozgató- és kevert agyidegek szerint csoportosíthatjuk.
3.9.1. Tisztán érző agyidegek
I. Szaglóideg (nervus olfactorius)
A felső orrkagylóban található primer érzékhámsejtek nyúlványai adják. Az érzékhámsejtek axonjai bejutnak a koponyába (lamina cribrosa területén) és a bulbus olfactoriusban végződnek, ahol más sejtekkel ún. glomerulusokat alkotnak. Az innen kiinduló másodlagos neuronok hosszú nyúlványai a tractus olfactoriust adják, amelyek az agykéreg alsó frontális részéhez, illetve a limbikus rendszer területeire projiciálnak.
II. Látóideg (nervus opticus)
A retinában található dúcsejtek axonjai alkotják az ideget, melyek a chiasma opticum területén féligátkereszteződnek (hemidecussatio). A kereszteződésük után, mint látópálya (tractus opticus) fut a talamusz CGL (corpus geniculatum laterale) magjához, és ott átkapcsolódnak. Az innen kiinduló rostok az elsődleges látókéregbe (Brodmann 17) futnak.
VIII. Halló- és egyensúlyozó ideg (nervus vestibulocochlearis)
Az agyideg rostjai egyrészt az egyensúlyozó szervből (lásd 5.3. fejezet), másrészt a Corti-féle szervből (lásd 5.2. fejezet) indulnak ki. Az innen jövő ingerületet a dúcsejtek (ganglion vestibulare és ganglion spirale) veszik át, melyek a koponya üregébe lépve a megfelelő magokhoz (pl. Deiters mag, nucleus cochlearis) futnak.
3.9.2. Tisztán motoros agyidegek
III. Közös szemmozgató ideg (nervus occulomotorius)
A középagyból induló ideg, mely a szem mozgásait biztosító finom szemizmokat idegzi be.
IV. Sodorideg (nervus trochlearis)
A sodorideg nagyon vékony, hosszú lefutású agyideg, mely az agytörzs hátsó oldalán lép ki. Kilépése után kereszteződik, majd előrefut a koponyagödörben és a szemüregbe lép. Az ideg a szemgolyó felső ferde izmát (m. obliquus superior) idegzi be.
VI. Távolító ideg (nervus abducens)
Az ideg motoros magja a nyúltvelőben található, innen kilépve fut a szemüregbe, ahol a felső szemgödri hasadékon keresztül lép be. A motoros ideg az oldalsó egyenes szemizmot idegzi be.
XI. Járulékos ideg (nervus accessorius)
A tisztán motoros ideg a nyúltvelőből lép ki, és feladata a trapéz- és fejbiccentő izmok, valamint a lágy szájpad, garat és gége izmainak beidegzése.
XII. Nyelv alatti ideg (nervus hypoglossus)
A nyúltvelőből eredő mozgató ideg a nyelv izomzatának mozgató beidegzésére, a nyelv alakjának változására szolgál.
3.9.3. Kevert agyidegek
V. Háromosztatú ideg (nervus trigeminus)
A kevert agyideg a fej legnagyobb részének érző idege, érzőrostjai részt vesz a fájdalom, hő, nyomás érzékelésében. Motoros rostjai egyebek között szerepet játszanak a rágóizmoknak a beidegzésében.
VII. Arcideg (nervus facialis)
Több funkcióval rendelkező kevert agyideg. A nyelv elülső kétharmadának specifikus érző idegellátását biztosítja. Beidegzi az arc mimikai izomzatát, a fül izmait, a platysmát. Paraszimpatikusan beidegzi az állkapocs alatti mirigyet (glandula submandibularis) és a nyelv alatti mirigyet (glandula sublingualis), valamint a könnymirigyet (glandula lacrimalis) egyaránt.
IX. Nyelv-garat ideg (nervus glossopharyngeus)
Fő motoros magja a nyúltvelőben helyezkedik el, ahol a bolygóideg (X. agyideg, nervus vagus) magjával közösen ered. Paraszimpatikus rostjai a fültőmirigyet(glandula parotis) idegzik be. Érzőrostjai általános érzéseket, valamint ízérzéseket közvetítenek. A közös fejverőér (arteria carotis communis) elágazódásánál található nyomásérzékelő receptorból jövő ingerületet is szállítja, így szerepet játszik az artériás vérnyomás szabályozásában,
X. Bolygóideg (nervus vagus)
A legnagyobb paraszimpatikus ideg, mely beidegzi a mellüreg és hasüreg szerveit egyaránt. Motoros magja, mely a nyúltvelő agytörzsi hálózatos állományában található, ellátja a garatfűző izmokat, és a gége belső izmait. Érzőingerületet a nyelőcső, a hörgők, valamint a szív felől szállít.
*Érdekesség: Kutatásokkal sikerült bizonyítani, hogy a Parkinson-kór egyik oka a bélrendszerre tehető, és ennek közvetítésében a bolygóideg szerepet játszhat. Kimutatták, hogy azoknál a betegeknél, akiknek a bolygóidegét a gyomorfekély kezelése érdekében átvágták (vagotómia), azok a páciensek védelmet nyertek a Parkinson-kór ellen.
3.10. A limbikus rendszer
3.10.1. A Papez-kör
1937-ben James Papez írt le először egy funkcionális kört, mely magában foglalja a nagyagykéreg belső felszínének, a kérgestest körül elhelyezkedő részeit, illetve a talamuszt és a hippokampuszt. Akkoriban ezt szaglóagynak nevezték el (rhinencephalon) és feltételezték, hogy a szaglóinformációk feldolgozásában játszik szerepet. Ma már tudjuk, hogy a Papez-kör és egyéb agyterületek együttesen, az ún. limbikus rendszert alkotva az érzelmeink legfontosabb agyi központjai. A Papez-körön kívül a limbikus rendszer részének tekinthetjük az amigdalát, a harag, düh és a félelem központját, valamint a hipotalamusz egyes régióit is.
A 3-10. ábrán nyomon követhetjük a Papez-körben az információáramlás irányát:
gyrus cinguli > hippokampusz > fornix > corpus mamillare > talamusz > gyrus cinguli
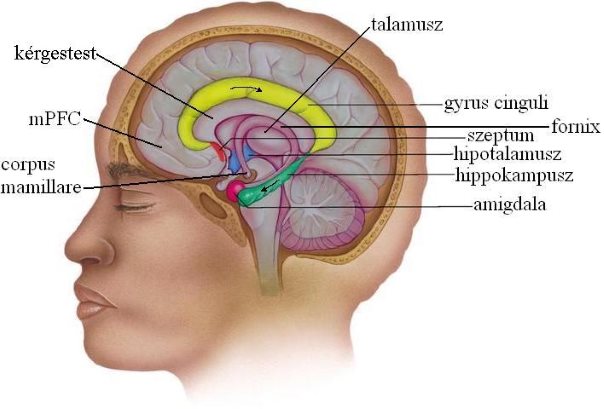
3-10. ábra: A limbikus rendszer a Papez-körrel
3.10.2. A „szociális agy” és az emberi evolúció
A prefrontális agykéreg középső része (mediális prefrontális kéreg, mPFC) és a limbikus rendszer, a szeptumon keresztül szoros kapcsolatban áll egymással (3-10. ábra). Kettejük működése együttesen nagy hatással van az ember szociális magatartására. A limbikus rendszeren belül az mPFC az amigdalával van kapcsolatban, kettejük együttes működését nevezzük „szociális agynak”. Az emberben különösen nagy méretűvé vált mPFC, gátló bemeneteket szolgáltatva a limbikus rendszer felé feltételezhetően szerepet játszik érzelmeink „kordában tartásában”. Egyes elméletek szerint, ez a kölcsönhatás tette lehetővé a társadalomalkotást az emberi evolúcióban, mivel lehetőséget adott az emberek közötti kooperációra azáltal, hogy az egymás közötti agressziót lecsökkentette. Valószínűsíthető az is, hogy az emberi evolúció egyik kulcsfontosságú lépése, a tűz megszelídítése is annak volt köszönhető, hogy a megnövekedett nagyságú mPFC le tudta gátolni az amigdalából érkező félelem-információkat. A félelmek legyőzésére az élővilágban szinte csak az ember képes. A fejlődő kooperatív magatartás, aztán az eszközhasználat és a kommunikáció fejlődéséhez vezetett, mely a nyelv kialakulását segítette elő.
*Kiegészítés: Egyes kórképek kialakulásának magyarázata lehet szintén az mPFC-amigdala kettősének eltorzult működése. Ha az mPFC válik dominánssá, akkor az értelmi intelligencia erősödhet, mialatt az érzelmi intelligencia gyengül, ez lehet az autizmus tüneteinek egyik indoka. Ezzel ellentétben, ha az amigdala válik dominánssá, akkor az érzelmi intelligencia fokozódik, az ilyen emberek nagy szociális érzékenységgel rendelkezhetnek. Annak ellenére, hogy esetleg értelmi intelligenciájuk nem éri el az átlagot. Szélsőséges esetben ilyen lehet a Williams-szindróma. Az ebben szenvedő gyerekek - „mindenki kedvencei” - könnyen alakítanak ki kapcsolatokat, nagyon nyitottak, mozgásszabályozásuk nem megfelelő, és kevesen tudnak közülük megtanulni írni vagy olvasni.
3.11. Vegetatív idegrendszer
A vegetatív idegrendszer belső szerveink működésének, központi idegrendszertől viszonylag független (autonóm) szabályozását végzi. Működése akaratlagosan kevéssé befolyásolható. A három fő szabályozó rendszer (4-1. ábra) közül, a szomatikus idegrendszernél (érzőműködések, mozgatóműködések) lassabb, a hormonrendszernél viszont gyorsabb regulációt végez. Egy vegetatív szabályozási ciklus ideje percekben, órákban mérhető. A vegetatív idegrostok lefutására általánosan jellemző, hogy a központi idegrendszerből kilépő axonok (preganglionáris rostok) a célszerv előtt, még egyszer átkapcsolnak valamely perifériás vegetatív dúcban. Az innen kiinduló (posztganglionáris) rostok idegzik be a belső szervet. A posztganglionáris rost mindig velőhüvely nélküli axon, melyen sok gyöngyfüzérszerű megvastagodás (varikozitás) helyezkedik el. A neurotranszmitterek ezekből ürülnek ki és viszonylag lassan, diffúzióval jutnak el a célszerv receptoraihoz, ahol serkentő vagy gátló hatást fejthetnek ki.
3.11.1. Paraszimpatikus idegrendszer
Anatómiailag a paraszimpatikus rostok az agytörzsből (III., VII., IX., X. agyidegek), ill. a gerincvelő keresztcsonti szakaszából, mint gerincvelői idegek lépnek ki. Mivel perifériás vegetatív dúcaik a gerincvelőtől távol, közvetlenül a beidegzett szerv előtt helyezkednek el, preganglionáris rostjaik hosszúak, behálózzák az egész testüreget (3-11. ábra). Jellemző ingerületátvivő anyaguk az acetilkolin (ACh), mely a vegetatív dúcok idegsejtjein nikotin-típusú- (nAChR), míg a célszerveken muszkarin típusú acetilkolin-receptorokhoz (mAChR) kötődik.
A paraszimpatikus idegrendszer akkor aktiválódik, ha a szervezet pihen, feltölti tartalékait, regenerálódik, leginkább az étkezés utáni ellazult állapottal példázható. Ennek megfelelően ilyenkor a sejtekre a felépítő anyagcsere folyamatok (asszimiláció) túlsúlya jellemző. A légzés- és szívfrekvencia lassul, a vérnyomás csökken, a bőrerek kitágulnak. A vér nagy része a belekbe áramlik, segítve ezzel a tápanyagok emésztését és felszívását. A pupilla szűkül. A magasabb rendű központi idegrendszeri funkciók (tanulás, figyelem) csökkent hatékonysággal működnek, az agyműködésre is a pihenő állapot jellemző, szinkronizáltabb idegsejt működéssel, valamint alacsonyabb frekvenciájú EEG-vel.
3.11.2. Szimpatikus idegrendszer
A szimpatikus preganglionáris rostok a gerincvelő háti, illetve ágyéki szakaszából lépnek ki, és a gerincvelőhöz közel elhelyezkedő szimpatikus dúcokban (para-és prevertebrális dúcok) kapcsolnak át, ezáltal rövidebbek, mint a paraszimpatikus preganglionáris axonok. Az innen kiinduló hosszabb posztganglionáris rostok szintén velőtlenek és varikozitásokban végződnek, a beidegzett szervek közelében. Jellemző neurotranszmitterük a noradrenalin. Beidegzés szempontjából kivételt képez a mellékvese velőállománya, mely közvetlen (preganglionáris) idegi bemenetet kap. Az ideg a vertebrális dúcokon áthalad ugyan, de ott nem szinaptizál, posztganglionáris rost nincs. Paraszimpatikus beidegzése nincs. A közvetlen beidegzés szerepe az információátadás gyorsítása, így vészreakciókban (támadás, menekülés, stressz) gyorsabban ürülhet a mellékvesevelőben termelődő adrenalin és a noradrenalin a vérbe, melynek segítségével az élőlény hamarabb reagálhat a fenyegető ingerre (3-11. ábra).
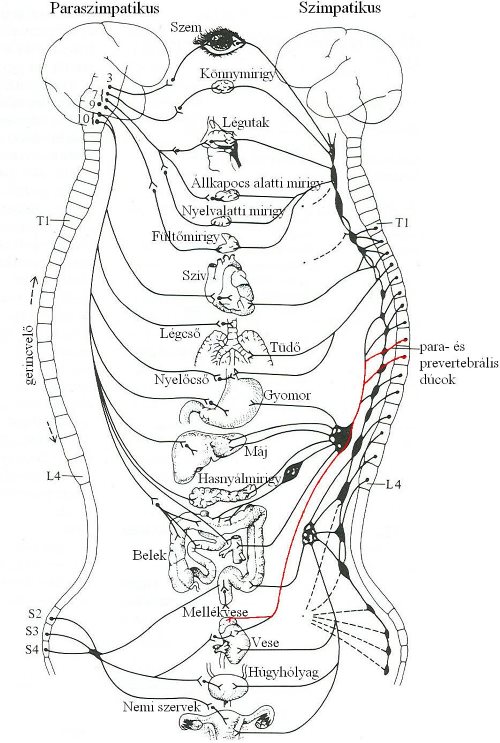
3-11. ábra: A vegetatív idegrendszer felépítése
Pirossal a mellékvesevelő direkt, szinapszis nélküli szimpatikus beidegzése látható
Rövidítések: T: thorakális (háti) szegmens, L: lumbális (ágyéki) szegmens, S: szakrális (keresztcsonti) szegmens
/Forrás: Withers, Comparative animal physiology, 1992./
Szimpatikus aktiváció általában akkor következik be, ha a szervezet felkészül valamilyen feladat megoldására, ha veszélyesnek ítélt szituációba kerül, vagy ha valamilyen stressznek van kitéve. Ilyenkor a sejteknek több energiára van szükségük, ezért a lebontó anyagcserefolyamatok (disszimiláció) aktiválódnak, melyek során a tartaléktápanyagok lebomlanak, nő a vércukorszint és a sejtekben ATP keletkezik. Több oxigénre van szükség, ezért nő a vérnyomás, fokozódik a szívritmus és a légzésfrekvencia. A bőrerek szűkülnek, míg a pupilla tágul. Mivel az emésztésre ilyenkor nem jut felesleges energia, a belek vérellátása és motorikája csökken, az emésztés gátolt. Az agy szimpatikus aktivációkor éberebb, az EEG-t magas frekvenciájú jelek jellemzik, bonyolult deszinkronizált neuronműködés figyelhető meg. A tanulási folyamatok és az emléknyomképződés hatékonysága rövidtávon javul, de krónikusan sokáig fennálló szimpatikus aktivációban (stressz) romolhat. A fiziológiás kézremegés, a vérnyomás emelkedés és az izomtónus erősödése miatt fokozódhat.
3.11.3. Szimpatikus vagy paraszimpatikus tónust fokozó szerek
A paraszimpatikus aktivátorok közé tartoznak az opiátok (ópium, morfin, heroin).
Szimpatikus aktivátorok: koffein (kávé), teofillin (tea), teobromin (kakaó, csokoládé), atropin (nadragulya), ide sorolhatók még az amfetamin-származékok (MDMA, ecstasy, speed) és a kokain, ill. az LSD és a meszkalin is.
A nikotin és az alkohol hatása összetettebb. A nikotin egyszerre viselkedhet paraszimpatikus és szimpatikus stimulánsként, míg a kis mennyiségű alkohol (pl. 1dl bor) paraszimpatikus hatást, a nagyobb mennyiségű alkohol pedig szimpatikus hatást vált ki. Mindezen szimpatikus vagy paraszimpatikus tónust befolyásoló szerek, nagymértékben képesek változtatni a szervezet mozgásszabályozó központjait.
| « Előző fejezet | Tartalomjegyzék | Következő fejezet » |
Események
Jelenleg nincs aktuális esemény.