TÁMOP-4.1.2.E-13/1/KONV-2013-0012
"Sporttudományi képzés fejlesztése
a Dunántúlon"
- -> Tananyagfejlesztés
- -> A vázizom...
- -> 4.












4.
4. AZ ERŐKIFEJTÉS MOLEKULÁRIS ÉS NEURÁLIS ASPEKTUSAI
- 4.1. Az izomrosttípusok
- 4.1.1. Tipizálás a rostok színe alapján: a történeti kiindulópont
- 4.1.2. Anyagcsere folyamatok szerinti rosttipizálás
- 4.1.3. Rosttipizálás miozin ATP-áz enzimaktivitás alapján (enzimhisztokémia)
- 4.1.4. Rosttipizálás miozin nehéz- és könnyűlánc típus alapján (immunhisztokémia)
- 4.1.5. Funkcionális és morfológiai különbségek az egyes rosttípusok között
- 4.2. A motoros egység
- 4.3. Az izom elektromos aktivitása
- 4.4. Az izom reflexmechanizmusai
A vázizom szerkezeti és működésbeli alapjainak elsajátítása után a következő két fejezetben az erő kifejtését befolyásoló tényezőkkel foglalkozunk. Az izomerő kifejtésének nagyságát, időtartamát és sebességét számos molekuláris, neurális, morfometriai, morfológiai és mechanikai faktor külön-külön, vagy akár együttes megjelenése befolyásolhatja, mely célorientált mozgásainkra hatással vannak. A tankönyv szerkesztésekor kézenfekvőnek tűnt, hogy az erőkifejtés morfometriai és morfológiai paramétereit (izomkeresztmet, izomhossz és rostelrendeződés) egy korábbi fejezetben ismertessük, az izom szerkezeti sajátosságainak tárgyalásával párhuzamban. Ebben a fejezetben az erőkifejtés neurális és molekuláris paramétereivel foglalkozunk a motoros egység definiálástól és az izomrosttípusok megismerésétől kezdve az izom reflexmechanizmusáig. Mivel az emberi mozgások minősége – függetlenül attól, hogy sportmozgás, vagy hétköznapi mozgás – ezeken a faktorokon múlik, ezért a következő fejezetek tartalma jelentős gyakorlati hasznosíthatósággal bír.
4.1. Az izomrosttípusok
4.1.1. Tipizálás a rostok színe alapján: a történeti kiindulópont
A kutatók már több mint száz éve foglalkoznak a különböző izomrosttípusokkal és már 1800-as években rájöttek arra, hogy egyes izomrostok eltérnek színben, anatómia szerkezetükben és funkciójukban. A legkorábbi megkülönböztetés Ranvier (1873) nevéhez fűződik, aki megfigyelte, hogy egyes izmok lassabban húzódnak össze és mindemellett jellegzetesen vörös színű, ami a magas (oxigén szállításért felelős) myoglobin koncentrációnak köszönhető. Később Denny-Brown (1929) megállapította, hogy egyes izmokban fehér és vörös színű rostokat is tartalmaz, és ezek keverten helyezkednek el. A kutató szerint a vörös rostok a testtartásért a fehér rostok pedig a gyors mozgásokért felelősek. A szín alapján történő rosttipizálás tekinthető tehát a legelsőnek.
4.1.2. Anyagcsere folyamatok szerinti rosttipizálás
Anyagcsere folyamatok alapján az egyes izomrostokat a szerint különböztetjük meg, hogy azok oxidatív vagy glikolítikus módon állítják-e elő a kontrakcióhoz szükséges ATP-t. E szerint a szempont szerint Peter és mtsai (1972) háromféle rostot különített el. Azokat a rostok, melyek magas oxidatív működést mutattak, lassú oxidatív (slow oxidative) rostoknak nevezték el (nemzetközileg elfogadott rövidítés az angol nyelvű terminusból: SO). Azokat a rostokat, amelyek magas glikolítikus működést mutattak, gyors glikolítikus (fast glycolitc) rostoknak nevezték (rövidítés: FG). Korábbi kutatásoknak köszönhetően az is bebizonyosodott, hogy nem mindegyik gyors rost fehér, ezért egy köztes rostot is meghatároztak a fent említett szerzők. A mind magas oxidatív, mind pedig magas glikolítikus tulajdonsággal bíró rostokat gyors oxidatív-glikolítikus (fast oxidative glycolitic) rostoknak nevezték el (rövidítés: FOG).
A magas oxidatív kapacitás azt jelenti, hogy a rostokban főleg aerob körülmények között, vagyis oxigén jelenlétében megy végre az ATP termelés. Ez oxidatív magas szintű oxidatív enzimfolyamatok eredménye. Az ATP termelés a mitokondriumokban zajlik, tehát a lassú rostok mitokondrium tartalma magas. Ezzel szemben a gyors rostokban a glikolítikus enzimek aktivitása magas, ami lehetővé teszi, hogy a szénhidrátok oxigén jelenléte nélkül, anaerob módon égjenek el az ATP előállításához. Ilyenkor a glükóz lebontása nem éri el a citromsav ciklust, csupán a glikolízisen megy keresztül, amely a citoplazmában megy végbe. És mivel ez utóbbi lényegesen gyorsabb folyamat, ezért a glikolítikus rostok gyors rángású, viszont fáradékony rostok. Az oxidációs ATP termelési folyamat már lényegesen több időt vesz igénybe, ezért az a lassú rostokra jellemző. Az izomrostok energiatermelési módja tehát egyértelműen utal a funkcióra is. A köztes rostok mindkét energiatermelésben magas szintet mutatnak, ezekre a rostokra van szüksége leginkább az olyan sportolóknak, akiknek a sportágában valamennyi kondicionális képesség megjelenik (erő, gyorsaság, állóképesség).
4.1.3. Rosttipizálás miozin ATP-áz enzimaktivitás alapján (enzimhisztokémia)
A fentiekben leírt felfedezések idején a kutatók nem rendelkeztek olyan laboratóriumi eszközökkel, hogy az egyes rostokat molekuláris szinten vizsgálják. A hisztokémiai eljárások kibontakozása áttörést jelentett, ugyanis lehetővé vált az egyes izomrostok festése és elekronmikroszkóp alatti tipizálása. A hisztokémia a szövetek elektronmikroszkópos vizsgálatát jelenti (histo = szövet). Az egyes szövetekből vékony metszetet készítenek, mely metszetben a szövetben található enzimek aktivitását képesek mérni a kutatók. A szövetet egy bizonyos szubsztráttal kezelik, melyre a vizsgált enzim reakcióba lép. A reakció következtében létrejövő elszíneződés alapján vizualizálni tudjuk az enzim aktivitását az adott szövetben.
Már a 60-as évek végén igazolták, hogy a miozin molekulának jelentősége van a kontrakció sebességében. Az érdeklődést a kutatókban nyilván az kelthette fel, hogy az erőcsapás, vagyis a kereszthidakban létrejövő mechanikai munka a miozinfejek elmozdulásának köszönhető. Az izomrost típusok elkülönítésére az egyes rostokat Engel (1962) miozin ATP-áz enzimaktivitás alapján festette meg (4.1. ábra), amely jelenleg is elfogadott és gyakran alkalmazott módszer. A biopszia segítségével kivett izommintákat 9.4 pH-val rendelkező oldatba helyezve a gyors rángású izomrostok (melyek egyébként magasabb ATP-áz enzimaktivitással rendelkeznek) sötétebb árnyalatban jelentek meg, mint a lassú izomrostok. A gyors izomrostokat II-es, a lassúakat pedig I-es típusú rostoknak nevezték el. Később Brooke és Kaiser (1970) a II-es rostoknál további altípusokat különített el: IIA, IIB és IIC. Különböző pH-val kezelve ezek a rostok ugyanis eltérő elszíneződéssel reagáltak. Kereszthíd ciklus során a miozin ATP-áz enzim felelős az ATP lebontásának sebességéért a miozin fejen. A kereszthidak kialakulásának és megszűnésének, vagyis az aktin és miozin molekulák egymáson történő elcsúszásának a sebességét tehát a miozin ATP-áz enzim befolyásolja. Minél nagyobb ennek az enzimnek az aktivitása, annál gyorsabban képes az izomrost rövidülni.
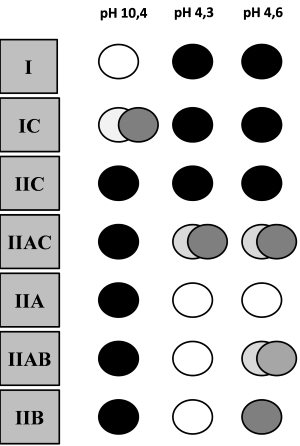
4.1. ábra. Izomrosttípusok meghatározása miozin ATP-áz enzimaktivitás alapján. A biopsziából nyert metszeteket különböző pH-val kezelik (10,4; 4,3 és 4,6), melyre az eltérő enzimaktivitással rendelkező rostok különböző színűre festődnek (Staron és Hikida 1992).
1992-ben Staron és Hikida egy olyan kutatómunkát publikált, amelyben a fentiekben ismertetett rosttípusokon kívül más típusokat is felfedezett. A kutatók egy 20 éves, erőedzésben jártas nő vastus lateralis izomzatából vett biopsziában szokatlan rostkompozíciókat találtak. Miozin ATP-áz enzimaktivitás alapján a C rostoknak három további altípusát különítette el: IC, IIC és IIAC rostokat. Az IC rost enzimaktivitása a I-es rostéhoz közelített, míg a IIAC rost enzimaktivitása a IIA rostéhoz közelített. Az IC rost mutatott némi gyors kontraktilitási hajlamot, a IIAC pedig némi lassú kontraktilitási hajlamot. Az eredmények azt mutatják, hogy az egyes rostok között nagyon finom átmenet alakult ki kontraktilitási sebességben, valamint oxidatív/glikolítikus tulajdonságban. Staron és Hikida alapján a 4.2. ábrán szemléltetjük az egyes rosttípusok sebességbeli és anyagcsere folyamatbeli sorrendiségét. Mindezek alapján a legnagyobb oxidatív kapacitással bíró és egyben a leglassabb rost az I-es típus. A IIB rost felé haladva egyre csökken a rostok oxidatív kapacitása, viszont nő a glikolítikus kapacitása és kontraktilitási sebessége. Az anyagcsere folyamatok alapján elmondható, hogy az I-es típusú rost a legkevésbé fáradékony, míg a IIB rost felé haladva egyre jobban hajlamosak a rostok a fáradásra. A lassú rostok inkább testtartásért, járásért, vagy egyéb lassú, hosszan tartó mozgásért felelnek. Ezzel szemben a gyors rostoknak a nagy sebességgel vagy nagy erővel végrehajtott mozgásokban van szerepe. Staron és Hikida nem zárták ki, hogy mivel a szomszédos rostok nagyon közel állnak egymáshoz a fenti tulajdonságok tekintetében, előfordulhat, hogy azok átalakulhatnak egymásba. Ezzel egy későbbi fejezetben foglalkozunk.
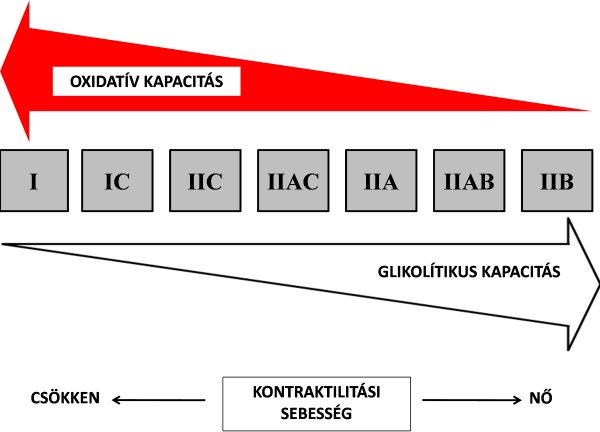
4.2. ábra. Miozin ATP-áz aktivitás alapján hisztokémiai úton elkülönített rosttípusok Staron és Hikida (1992) szerint. Az I-es rosttípustól haladva a IIB felé a köztes rostoknak nő a glikolítikus tulajdonsága és a kontraktilitási sebessége. Ellenkező irányban nő a rostok oxidatív tulajdonsága és csökken a kontraktilitási sebessége.
4.1.4. Rosttipizálás miozin nehéz- és könnyűlánc típus alapján (immunhisztokémia)
Az enzimhisztokémiai eljárások után a miozin molekula precízebb vizsgálatára az immunhisztokémia adott lehetőséget. A kutatók olyan antitesteket hoztak létre, melyekkel a miozin nehézláncnak is elkülöníthető változatait (izoformot) fedezték fel (Shiaffino és mtsai, 1989). 2. fejezetben már ismertettük, hogy a miozin (ha a két molekulát együtt nézzük) két nehéz (heavy chain) és négy könnyű láncból (light chain) tevődik össze. Az immunhisztokémiai vizsgálatok alapján három fő miozin nehézlánc típust különböztetünk meg: I, IIa és IIb. A kontrakció során az erőcsapás sebességét és erejét a miozin molekula nehéz lánc tartalma jelentősen befolyásolja (Aagaard és Andersen, 1998). A IIb izoformot tartalmazó miozin kereszthídjainak az erőkifejtése a leggyorsabb, az I-es izoformmal rendelkező miozin pedig a leglassabb. Fontos megjegyeznünk, hogy egy adott izomrost teljes hosszában ugyanaz a nehézlánc izoform található meg, tehát a nehézlánc alapján is tipizálhatjuk a rostokat.
A későbbi kutatásokban azt is igazolták, hogy a miozin nehézláncon található könnyűláncok is befolyásolják a kontrakció sebességét. VanBuren és mtsai (1994) egy kísérlet során eltávolították a könnyű láncokat és azt tapasztalták, hogy az aktin mentén a miozin elcsúszásának sebessége 50-80%-kal csökkent. Amint a könnyűláncokat visszahelyezték, a két filamentum mozgása egymás mellett visszanyerte eredeti sebességét. Minden egyes miozin nehézláncon egy esszenciális (essential light chain) és egy szabályozó (regulatory light chain) könnyűlánc található. Lowey és mtsai (1993) bizonyították, hogy ezek közül bármelyik eltávolítása jelentősen csökkenti a kontrakció sebességét. Elmondható tehát, hogy a miozin könnyűlánc bár nem vesz részt az ATP hidrolízisében úgy, mint a nehézlánc, mégis fontos szerepet tölt be a kontrakció sebességének szabályozásában. Az esszenciális könnyűláncnak négy típusa van, melyeket LC1s, LC1f, LC3s és LC3f rövidítést kaptak (megjegyzés: LC = light chain, s = slow, f = fast; a rövidítések a típusra és a sebességre is utalnak; pl.: LC1s lassabb, mint LC1f). A szabályozó könnyűláncnak két típusa van: LC2s és LC2f. Az izomrostok rövidülési sebességét a miozin molekula nehéz- és könnyűlánc alkotórészek határozzák meg. És mivel a könnyűláncnak ennyi változata van, és ezek többféleképpen variálódhatnak egy miozinon belül, érthető, hogy a (korábban ismertetett) ATP-áz enzimaktivitás alapján miért tudtak hétféle rosttípust elkülöníteni. A 4.1 táblázatban látható, hogy a fő rosttípusok milyen könnyűlánc variációkból állhatnak.
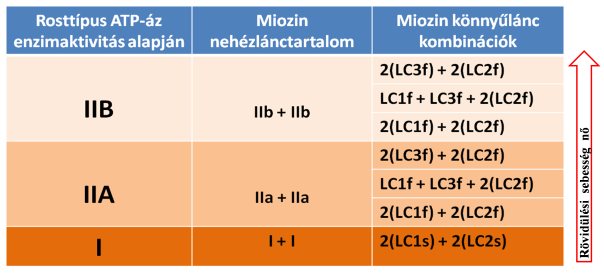
4.1. táblázat. Egy miozin molekula nehéz- és könnyűlánc összetevői az egyes rosttípusoknál. A teljes miozin molekula két nehézláncból
és négy könnyűláncból áll.
Megjegyzés: LC = könnyűlánc, f = gyors, s = lassú.
4.1.5. Funkcionális és morfológiai különbségek az egyes rosttípusok között
Az egyes izomrosttípusok a korábban említett miozin szerkezeten és enzimaktivitáson kívül számos más tulajdonságban is különböznek. A fő izomrosttípusok szerkezeti, élettani és funkcionális különbségeit a 4.2. táblázatban foglaltuk össze. Gyakorlati szempontból talán a leglényegesebb különbség a rövidülési sebességben és az erőkifejtésben van. A 2B típusú izomrost a leggyorsabban, az I-es típusú pedig a leglassabban rövidül, a sebességbeli különbségeket pedig az ismert miozin nehéz- és könnyűlánc kombinációk adják. Érdekes az a megállapítás, mely szerint a gyors rostok tubuláris rendszere (T tubulus és szarkoplazmatikus retikulum) sűrűbb a lassú rostokéhoz képest, amely arra utal, hogy a motoneuronokból érkező ingerület ugyanannyi idő alatt több miofibrillumhoz eljut, vagyis a teljes izom gyorsabban aktiválódik (Eisenberg, 1983).
Általánosságban elmondható, hogy az I-es típushoz közelebb álló (lassabb) rostok nagyobb oxidatív kapacitással bírnak, így állóképességük jobb, a fáradásnak jobban ellenállnak. A magas oxidatív kapacitás a magas oxidatív enzimkoncentrációnak köszönhető. A IIB-hez közelebb álló rostok glikolítikus kapacitása nagyobb, ezek fáradékonyabb rostok, kisebb az állóképességük. Ezekben a rostokban a glikolítikus enzimek koncentrációja magasabb. A IIA rost talán a legkedvezőbb azon sportolók számára, akik nagy sebességű mozgásokat hajtanak végre többször egymás után, hosszú ideig (pl. labdajátékosok). A IIA rostok egyaránt magas oxidatív és magas glikolítikus kapacitással bírnak. Az állóképességbeli különbség a mitokondriumok sűrűségének is köszönhető, mely magasabb a lassú rostokban.
A kutatók megállapították, hogy a főleg gyors izomrostokat tartalmazó izmok nagyobb erőkifejtésre képesek, szemben a főleg lassú rostokból álló izmokkal. A gyors izmokra jellemző, hogy megközelítőleg 22 N/cm2 erőt fejtenek ki, míg a lassú izmok csupán 10-15 N/cm2-t (lásd: Lieber, 2002). Bodine és mtsai (1987) azonban kimutatták, hogy a gyors rostok csupán kismértékben fejtenek ki nagyobb erőt a lassúakhoz képest. Ezt alátámasztja az a megállapítás, hogy a kétféle rosttípusban az egy szarkomerre eső kereszthidak száma azonos (Zappe és Maeda, 1985). A gyors izmok nagy erőkifejtő képességét a rostok beidegzési tulajdonságában kell keresni.
A rostok szarkomerszerkezetének tekintetében jelentős különbség található a Z-vonalak vastagságában. A gyors rostok szarkomereiben vékonyabbak a Z-vonalak, mint a lassú rostokéban. Az M-régióban is jelentős különbség van, mely szintén vékonyabb a gyors rostokban. Végül meg kell említenünk, hogy a gyors rostok vastagabbak, mint a lassú rostok.
A lassú és gyors rostok keverten, mozaikszerűen oszalanak el egy izmon belül, de megoszlási aránya egyénenként és izmonként változó. A rosttípusok eloszlása utalhat az adott izom funkciójára. Armstrong és mtsai (1982) kutyák izomainak vizsgálatában igazolta, hogy elsősorban az antigravitációs, tehát a testtartásért felelős extenzor izmokban a legmagasabb az I-es rostok aránya. Megállapították azt is, hogy egy azonos izmon belül a mélyebb izomrétegekben nagyobb volt az I-es rostok aránya, mint a felszíni rétegekben. Embreknél az antigravitációs izmokban általában kb. 50-50% a lassú és gyors rostok aránya, de a soleus izomban 80% (Gollnick és mtsai, 1974), a tibialis anteriorban pedig 73% (Johnson és mtsai, 1973) is lehet a lassú rostok aránya. Ugyanakkor a szemhéj zárásáért felelős orbicularis oculi izomban mindössze 15% az I-es rostok aránya (Johnson és mtsai, 1973).
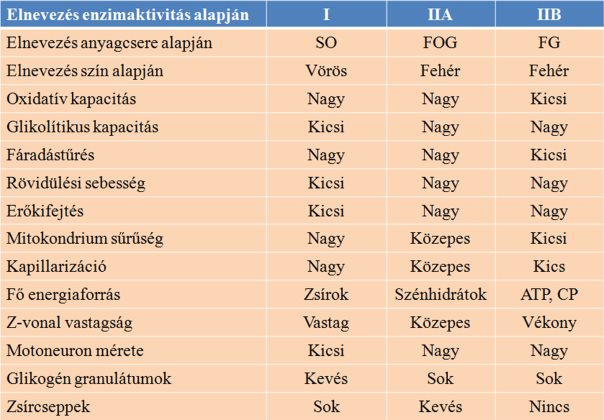
4.2. táblázat. A fő izomrosttípusok közötti élettani, szerkezeti és funkcionális különbségek. SO = slow oxidatív, FOG = fast oxidative glycolitic, FG = fast glycolitic, ATP = adenozin trifoszfát, CP = kreatinfoszfát.
4.2. A motoros egység
Idáig az izomrostok szerkezeti, élettani és funkcionális tulajdonságaival foglalkoztunk, de működésük kizárólag idegrendszeri összeköttetéssel biztosított. A következőkben arról olvashatunk, hogy milyen idegrendszeri működési mechanizmuson keresztül szabályozhatjuk mozgáskoordinációnkat és erőkifejtésünk mértékét.
Az egyes izomrostokat a gerincvelőben található motoros idegek (α motoneuronok) nyúlványai (axonjai) idegzik be. Az ingerület ezeken az axonokon keresztül jut el a rostokig. Egy α motoneuron és az általa beidegzett izomrostokat együttesen motoros egységnek (motor unit) nevezzük (4.3. ábra). Az α motoneuronok a gerincvelő elülső szarvában találhatók. Minden egyes motoneuronból egy axon indul el az izmok felé, de amint az axon eléri az izmot, szerteágazik, és mindegyik ág egyetlen izomrosthoz fut be. Egy motoneuron így több izomrostot is beidegez, és ez azt jelenti, hogy ha ingerület érkezik, akkor az valamennyi beidegzett rosthoz eljut. Azonos motoros egységhez tartozó izomrostok közül tehát vagy mindegyik egyszerre aktiválódik, vagy ingerület hiányban egyik sem („mindent vagy semmit” törvény). Egy izomban sok motoros egység található, és ezek száma izmonként rendkívül változatos. A motoros egységekhez tartozó rostszám is változó lehet. Egy adott izom erőkifejtését az határozza meg, hogy egyidejűleg hány motoros egység kapcsolódik be.
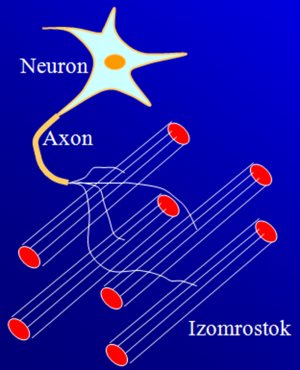
4.3. ábra. A motoros egység összetevőinek sematikus ábrája.
4.2.1. A motoros egység típusai
Burke (1967) alapján a motoros egységeknek három típusát különítjük el: lassú motoros egység (slow motor unit; nemzetközi jelölés: S), gyors fáradástűrő motoros egység (fast fatigue resistante motor unit; nemzetközi jelölés: FR) és gyors fáradékony motoros egység (fast fatiguable motor unit; nemzetközi rövidítés: FF). A motoros egységek leggyakoribb megkülönböztetése a fáradási index alapján történik. A fáradási indexet úgy állapítjuk meg, hogy ugyanakkora frekvenciájú folyamatos stimuláció alatt vizsgáljuk, hogy a különböző motoros egységek erőkifejtése mennyivel csökken adott idő alatt (mekkora fáradás következik be). Lassú motoros egység 2 percig tartó 40 Hz-es stimulációjánál a motoros egységhez tartozó izomrostok erőkifejtése nem csökken a kiindulási érték 75%-a alá. Ugyanennyi idő alatt azonban a gyors fáradékony motoros egységek erőkifejtése a kiindulási erőszinthez képest 25% alá esik. A köztes, gyors fáradástűrő motoros egység pedig a kettő között, tehát 25% és 75% közötti értékre csökken.
Burke (1967) megállapította, hogy a különböző típusú motoros egységek egymástól eltérő rostokat tartalmaznak: az FF (gyors fáradékony) típusú motoros egységek FG típusú (gyors glikolítikus) rostokból állnak, az S típusú (lassú) motoros egységek pedig SO típusú (lassú oxidatív) rostokból. Értelemszerűen, az FR típusú (gyors fáradástűrő) motoros egységet FOG típusú (gyors oxidatív-glikolítikus) rostok alkotják (4.4. ábra). Fontos megjegyeznünk azonban, hogy bár egy adott motoros egység ugyanolyan típusú rostokból áll (a metabolikus tulajdonságot figyelembe véve), számos rost valamelyest mégis különbözhet azonos egységen belül (lásd az előző fejezetben: miozin ATP-áz aktivitás alapján) (Martin és mtsai, 1988). Érdekesség még az is, hogy a motoros egységek típusát (nevezetesen az egységekhez tartozó rostok típusát) a hozzá tartozó α motoneuron határozza meg. Ennek bizonyítását a klasszikus keresztinnervációs állatkísérlet eredményével magyarázhatjuk. Amikor a lassú és a gyors motoros egység axonjait felcserélték, a lassú rostok gyorssá, a gyors rostok pedig lassúvá alakultak át.
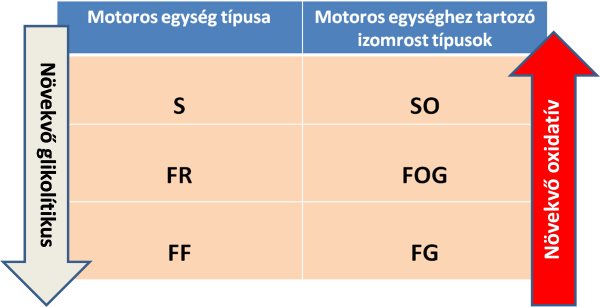
4.4. ábra. Motoros egység típusok és a hozzá tartozó izomrostok típusai metabolikus tulajdonság alapján tipizálva. S = slow oxidative (lassú oxidatív), FR = fast fatigue resistant (gyors fáradástűrő), FF = fast fatiguable (gyors fáradékony). A nyilak iránya növekvő tendenciát jelent.
Bodine és mtsai (1987) megfigyelték, hogy a gyors motoros egységek nagyobb erőt képesek kifejteni, mint a lassúak. Mindezt azzal magyarázták, hogy a gyors egységekhez több izomrost tartozik, és ezek a rostok vastagabbak is (vagyis több miofibrillum találahtó benne) (4.5. ábra). Mivel egy lassú és egy gyors izomrost egységnyi keresztmetszetre eső erőkifejtése valójában nem, vagy csak minimális mértékben különbözi, ezért valószínűsíthető, hogy a gyors motoros egységek valóban a nagyobb rostszám miatt erősebbek.
4.2.2. A motoros egység, mint az erőkifejtés funkcionális egysége
Az előző fejezetben már említettük, hogy a központi idegrendszer az akciós potenciálok impulzusainak sűrűségével képes az erőkifejtés mértékének finom szabályozására. A különböző méretű és típusú motoros egységek bekapcsolásának variálásával további szabályozási lehetőség áll rendelkezésünkre az erő kifejtésekor. A motoros egység a teljes izom erőkifejtésének funkcionális egysége. Egy izom pillanatnyi erőkifejtését részben az határozza meg, hogy az izomban a létező összes motoros egységből (motor unit pool) mennyi aktiválódik egyidejűleg. Mivel az erőkifejtést az aktív kereszthidak száma határozza meg, érthető tehát, hogy minél több motoros egység kapcsolódik be egyszerre, annál nagyobb lesz az izom erőkifejtésének mértéke. Nagyon nagy erő kifejtésekor a motoros egység készletnek akár 80-90%-a is bekapcsolódhat. 100%-os kihasználás nem létezik, bár a pszichés állapot jelentősen növelheti a bekapcsolt motoros egységek számát. Magasan motivált állapotban, vagy életveszélyes helyzetben az ember szokatlanul nagy erőkifejtésre képes. Az edzettségi állapot is jelentősen befolyásolja a motoros egységek kihasználásának képességét. Tegyük fel, hogy egy edzetlen és egy edzett ember azonos izomtömeggel rendelkezik. Szinte bizonyos, hogy az azonos izomtömeg ellenére az edzett ember jóval nagyobb erőkifejtésre képes, mint az edzetlen csupán azért, mert izomzatának nagy arányát képes egyszerre működtetni. A súlyemelők tipikusan olyan csoportját képezik az embereknek, akik kis testtömegük és izomtömegük ellenére rendkívül nagy tömegű súlyokat képesek emelni. A súlycsoportos sportágak más szereplői is, mint például a birkózók, cselgáncsozók, vagy ökölvívók edzésprogramja is arra irányul, hogy a sportolók izomerejét a végletekig fokozzák a testtömeg gyarapodása nélkül. A motoros egységek egyidejű bekapcsolását/kihasználását (motor unit recruitment), mint az izom egyik fontos neuromechanikai tulajdonságát, motoros egység szinkronizációnak (motor unit synchronization), vagy más néven intramuszkuláris koordinációnak (intramuscular coordination) nevezzük. Az izomnak ezt a tulajdonságát értelemszerűen a szinkronizációs erőfejlesztési módszerrel lehet fejleszteni, melynek lényege az, hogy a maximális erőkifejtéshez képest nagy ellenállást (90-100%) és kevés ismétlésszámot (1-5) alkalmazunk az erőfejlesztő edzés során. Az excentrikus kontrakciókkal végzett edzésnél a legyőzhető ellenállás maximumának 150%-val végzett gyakorlatokat javasolják, amely kiválóan javítja az intramuszkuláris koordinációt.
Az izom erőkifejtésének fokozására az aktív motoros egységek számának növelése meleltt van egy másik mód is, a motoros egységek kisülési frekvenciájának növelése. Egy adott motoros egységhez tartozó rostok akkor fognak nagyobb erővel összehúzódni, ha nagyobb frekvenciájú impulzusok érkeznek hozzájuk a motoros neurontól. A jelenséget frekvencia modulációnak nevezzük. Erőkifejtésünket tehát két úton növelhetjük: több és több motoros egység bekapcsolásával, és a már bekapcsolt motoros egységek kisülési frekvenciájának növelésével.
Köztudott, hogy amennyiben kis erőkifejtésre van szükségünk célorientált mozgásunk elvégzéséhez, kevesebb motoros egység aktiválódik. Egy kisebb tömegű tárgy megemeléséhez nincs szükség annyi kereszthíd egyidejű kapcsolódására, mint a súlyemelők esetében a nagy tömegű súlyok emelésénél. Egy célorientált mozgás precíz kivitelezésekor a feladat elvégzéséhez szükséges erő függvényében kell a motoros egységeket aktivizálnunk. Egy pohár víz megemelése kis erőkifejtést igényel, tehát a mozgásban résztvevő izmoknak csak kevés hányadát működtetjük. Ha a szükségesnél több motoros egység is bekapcsolódik, túlságosan nagy erőt fejtünk ki, a vizet kiboríthatjuk a pohárból. Az célorientált mozgásoknál nemcsak az egy izmon belüli motoros egységek szinkronizációjának variálásával szabályozzuk mozgásunkat, hanem több különböző izom egyidejű aktiválásával vagy relaxálásával. A fentiekben említett példánknál, a pohár víz megemelésében több izom is részt vesz. Ezeknek az izmoknak a szinkronitása vagy bekapcsolásuk időbeli sorrendisége jelentősen befolyásolja a mozgás végeredményét. Két vagy több különálló izom működésének összhangját, szinkronitását, bekapcsolási sorrendiségét intermuszkuláris koordinációnak (intermuscular coordination) nevezzük. Az intermuszkuláris és az intramuszkuláris koordináció együttesen szabályozza a mozgások kimenetelét. Míg az előbbi az izmok bekapcsolási sorrendiségét, az utóbbi az egyes izmok erőkifejtését szabályozza.
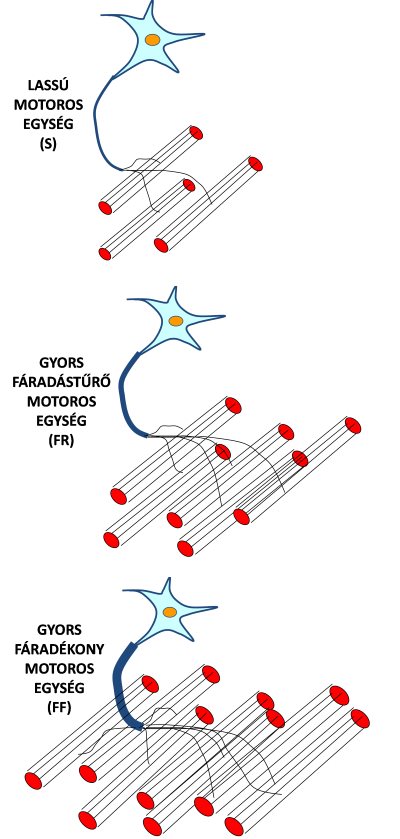
4.5. ábra. A motoros egység típusok fő anatómiai különbségei. Minél gyorsabb egy motoros egység, annál nagyobb a hozzá tartozó α motoneuron vastagsága, valamint a beidegzett izomrostok vastagsága és száma. S = slow oxidative (lassú oxidatív), FR = fast fatigue resistant (gyors fáradástűrő), FF = fast fatiguable (gyors fáradékony).
4.2.3. A motoros egységek bekapcsolási mechanizmusa az erőkifejtés szabályozására
Minél több motoros egység kapcsolódik be egyidejűleg, annál nagyobb tehát a teljes izom erőkifejtése. De vajon milyen szerepük van az egyes motoros egység típusoknak? Vajon milyen úton kapcsolódnak be azok különböző célfeladatoknál? Henneman és mtsai (1965) vizsgálatukban megfigyelték, hogy azok a motoros egységek, amelyek nagyobb erőt fejtettek ki, a hozzá tartozó axonon érkező elektromos impulzusok amplitúdója nagyobb volt, míg a kisebb erőkifejtést produkáló motoros egységeknél kisebb amplitúdójú volt az impulzus. Az impulzusok nagysága pedig függ az axon keresztmetszetétől. Vastagabb axonon nagyobb elektromos impulzus halad végig. A gyors motoros egységekhez vastagabb axon és sok izomrost tartozik, a lassú egységekhez pedig vékonyabb axon és kevesebb izomrost (4.5. ábra). A gyors motoros egységek nagy ereje még abból is adódik, hogy gyors rostokat tartalmaznak, melyek vastagabbak, vagyis több miofibrillum van bennük. Végül fontos megemlíteni azt is, hogy a gyors motoros egységek ingerlési küszöbe (recruiting threshold) magas, a lassúaké alacsony. Ingerlési küszöb alatt azt a minimális erősségű idegi impulzust értjük, amely az izomrostokban kontrakciót vált ki. Edzett személyek azért is képesek nagyobb erőkifejtésre, mert a magas ingerlési küszöbbel rendelkező motoros egységeket/izomrostokat is képesek bekapcsolni, szemben az edzetlen emberekkel. Ráadásul erőedzések ismétlésével a magas ingerküszöbű motoros egységek/izomrostok átalakulhatnak alacsonyabb ingerküszöbűvé. Ezzel a témával egy későbbi fejezetben foglalkozunk, amikor az izom alkalmazkodóképességét tárgyaljuk. Az alábbiakban részletezzük, hogy különböző erőkifejtési szituációkban az egyes motoros egység típusok milyen bekapcsolási sorrendet mutatnak.
- Amikor erőkifejtésünk lehetséges maximumát fokozatosan kívánjuk elérni, akkor először az S, majd az FR, végül pedig az FF motoros egységek fognak bekapcsolódni (4.6. ábra). Vagyis először a kis motoros egységek (kevés rostszámmal és vékony axonnal) kapcsolódnak be, majd az erőkifejtést fokozatosan növelve egyre nagyobb egységek kapcsolódnak be. Ezt a jelenséget „méretelvnek” (size principle) nevezzük (Henneman és mtsai, 1965). Képzeljük el, hogy egy szorítóerő-mérőt fokozatosan, egyre nagyobb erővel szorítunk. A szorítás elején elegendő a lassú egységek erőkifejtése is, de egyre erősebb szorítás esetén a gyors egységek ereje hozzáadódik a lassúakéhoz (összegződés).
- Amennyiben hirtelen szeretnénk nagy erőt kifejteni, akkor az eltelt idő rövidsége miatt csak a gyors motoros egységek fognak bekapcsolódni az erőkifejtésbe, a lassúak nem lesz idejük. Egy teljes sebességgel végzett ugró, dobó, vagy rúgó mozgásnál például a gyors motoros egységek és a hozzá tartozó gyors rostok szelektíven kapcsolódnak be. A nagy sebességgel végzett edzésgyakorlatoknak óriási jelentőségük van az olyan sportágakban, amelyekben a gyorsaság, a gyorserő, vagy a robbanékonyerő fontos. Ezekkel a gyakorlatokkal ugyanis a gyors motoros egységeket és hozzá tartozó gyors izomrostokat stimuláljuk.
- Amennyiben nagy erőkifejtést hosszú ideig próbálunk meg fenntartani (például maximális izometriás erőkifejtés 20-30 mp-ig), akkor egy bizonyos idő után az erő fokozatosan csökken, amit fáradásnak nevezünk. Ebben az esetben először a gyors motoros egységek fognak kikapcsolni, hiszen ezek a fáradékonyabbak, majd később a lassúak is.
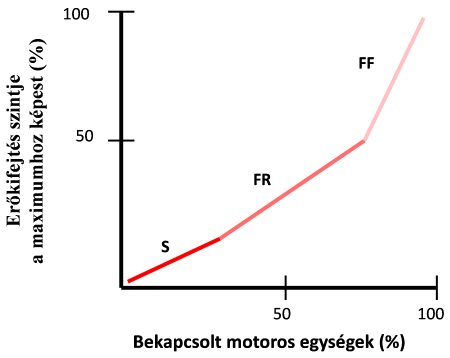
4.6. ábra. Motoros egységek bekapcsolási sorrendje fokozatosan növekvő, maximumig történő erőkifejtés esetén („méretelv”). Megjegyzés: a vízszintes tengelyen az akaratlagosan bekapcsolható motoros egységek maximális száma jelképezi a 100%-ot. S = slow oxidative (lassú oxidatív egység), FR = fast fatigue resistant (gyors fáradástűrő egység), FF = fast fatiguable (gyors fáradékony egység).
4.3. Az izom elektromos aktivitása
Emlékezzünk vissza a korábbi fejezetekre, amikor ismertettük a vázizom egyik lényeges tulajdonságát, az ingerelhetőségét. Megtárgyaltuk, hogy a központi idegrendszer neuronjaiból érkező elektromos impulzusok (akciós potenciálok) sokasága éri el a vázizmot, mely szétterjed az izomrostok membránján és kontrakciót hoz létre. Az izomnak ezt az elektromos jelenségét évtizedek óta vizsgálják a tudósok. Az izom elektromos aktivitását vizsgáló eszközt elektromiográfnak, a vizsgálati módszert elektromiográfiának (EMG) nevezzük.
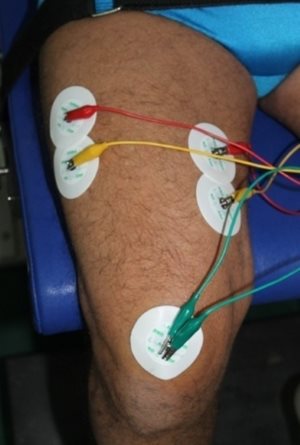
4.7. ábra. Elektromiográfia mérése felületi elektródák segítségével. A quadriceps izom erőkifejtésekor a vastus lateralis és medialis izmok felszínéről elvezetett feszültségjelek kerülnek elemzésre.
Az elektromiográfia nem közvetlenül az izmok erőkifejtésének kapacitását méri, hanem az azzal összefüggő idegi tevékenységet vizsgálja. Az idegi impulzusok erősségétől és sűrűségétől függ az izmok erőkifejtésének mértéke. Az EMG az izom elektromos aktivitását méri, vagyis azoknak az idegi impulzusoknak az összességét, amelyek az izmot elérik. A sporttudományban az EMG méréseket rendkívül jól vezető felületi elektródák segítségével végzik (4.7. ábra). Az elektródák az izom felszínén érzékelt igen alacsony feszültséget (mikro Volt nagyságrendű) egy erősítő felé vezetik, és a felerősített jeleket egy képernyőn jelenítjük meg, melyet elektromiogrammnak nevezünk (4.8. ábra). Ezt a nyers EMG jelet (raw EMG signal) számos matematikai műveletet követően (szűrés, simítás, rektifikálás) (4.9. ábra) számszerűsíthető adattá alakítjuk. Az elektromiogrammon ugyanazok a nagyságú hullámok azonos időintervallumokban ismétlődnek. A szűrés folyamata azt jelenti, hogy a nem vázizomra jellemző frekvenciájú hullámokat kivágjuk az elektromiogrammból (ilyen idegen hullám például a hálózati feszültség 50Hz frekvenciújú hulláma, amely megjelenhet az elektromiogrammunkon). A rektifikálás (négyzetre emelés) azt jelenti, hogy az elektromiogramm minden egyes pontját négyzetre emeljük, és ezzel csak pozitív értékeket kapunk. A simítás pedig azt jelenti, hogy a görbén több egymás utáni pont értékét átlagoljuk, így a görbén található durvább kiugrások kiegyenlítődnek.
Az egyik leggyakrabban alkalmazott adatfeldolgozási módszer az, amikor az EMG görbe alatti területet számoljuk ki (integrált EMG) (Váczi és mtsai 2011). Minél nagyobb a görbe alatti terület, annál erősebb idegi impulzusok érkeznek az izomhoz, amely általában nagyobb erőkifejtést is jelent.
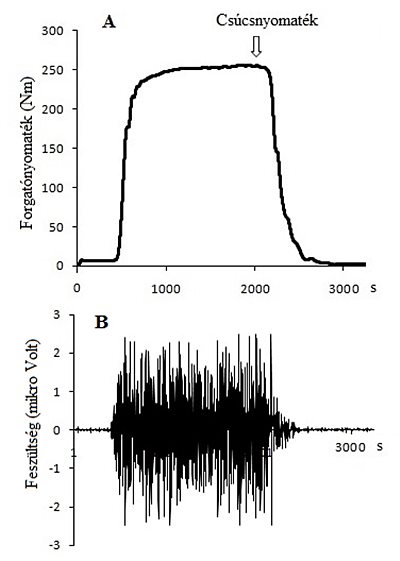
4.8. ábra. Térdextensio közben regisztrált forgatónyomaték-idő görbe (1000 Hz mintavételi frekvencia) maximális akaratlagos izometriás erőkifejtés alatt Multicont II típusú dinamométeren (A). Az erőkifejtés közben regisztrált (1000 Hz mintavételi frekvencia), a dinamométerrel időben szinkronizált nyers EMG jel a vastus lateralis izom elektromos aktivitását mutatja (B).
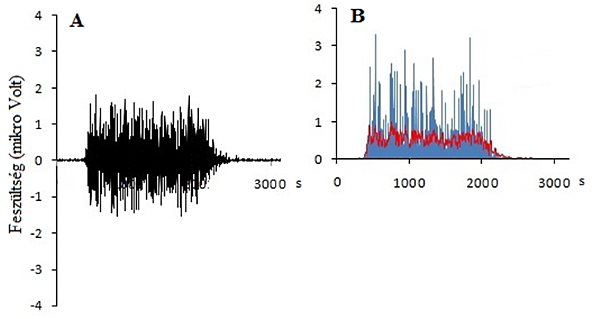
4.9. ábra. Nyers EMG jel (A) feldolgozása (1000 Hz mintavételi frekvencia). A nyers adatsor minden adatának négyzetre emelése (rektifikálása; kék görbe) és simítása (piros görbe) lehetővé teszi, hogy az EMG jelet kvantitatívan elemezhessük. Ennél a példánál a piros görbe alatti terület kiszámolásával kapjuk meg az integrált EMG (iEMG) értékét, mely az erőkifejtés során bekapcsolt izomrostok szinkronizált működéséről ad információt.
Az EMG vizsgálatok előnye az, hogy az izom morfometriai (pl. keresztmetszet) és metabolikus (anyagcsere) folyamatai kizárva tudjuk vizsgálni az erőkifejtésért felelős idegi mechanizmusokat. Meg tudjuk becsülni például, hogy egy edzésprogram mikor és milyen arányban okozott neurális adaptációt.
Az EMG-t használják még a fáradás vizsgálatára is. Amennyiben csökken az EMG jel erőssége, az a központi idegrendszer fáradását jelzi. Az izom fáradásáról részletesebben írunk egy későbbi fejezetben.
Az EMG vizsgálatokat huzalos elektródákkal is lehet végezni. Egy vékony huzalt vezetnek be az izom mélyére egy tű segítségével, amely elektródaként szolgál. Így a huzalos elektródával képesek vagyunk akár egyetlen motoros egység aktivitását is vizsgálni. Ilyen mérési eljárást inkább a tudományos alapkutatásokban és a klinikai vizsgálatokban alkalmaznak.
4.4. Az izom reflexmechanizmusai
Tankönyvünkben eddig az akaratlagos kontrakciókkal foglalkoztunk, de figyelembe kell vennünk, hogy számos mozgás során akaratlanul is működésbe lépnek izmaink. Előfordulhatnak olyan mozgásszituációk, amikor olyan gyorsan kellene cselekednünk, hogy azt akaratlagosan nem tudnánk megtenni. Ilyenkor a vázizom reflexszerű működésre is képes. Az izom reflexműködései nem érintik a központi idegrendszer agykérgi részét, csupán a gerincvelőit, ezért a reflexet kiváltó inger és az arra bekövetkező válasz közötti eltelt idő (reflexidő) rendkívül rövid.
4.4.1. Az izomorsó működése: a nyújtási reflex
Számos reflexműködése van az izomnak (pl. fájdalomérző reflex, bőrérzékelési reflex) de mozgás- és sporttudományi szempontból az izom leggyakrabban vizsgált reflexe a nyújtási reflex (stretch reflex), vagy miotatikus reflex (myotatic reflex). A jelenség legegyszerűbben a patella reflexszel vizsgálható. Az ülő helyzetben lévő személytől azt kérjük, hogy a quadriceps izmot lazítsa el, majd egy kalapáccsal finoman a patella ínra ütünk. Az ütés után néhány ms elteltével a lábszár előre mozdul. De lássuk, hogy vajon milyen mechanizmuson keresztül valósul meg a nyújtási reflex.
A nyújtási reflex kétségkívül a legrövidebb idő alatt lezajló ingerlés-kontrakciós jelenség az izomban, hiszen monoszinaptikus kapcsolaton megy keresztül az ingerület. A reflex kiindulópontja az úgynevezett izomorsó (muscle spindle). Az izomorsók olyan feszülésérzékeny receptorok, melyek az izom hosszváltozását érzékelik. Nevéből adódóan orsó alakú, 80-250 mikrométer vastagságú képződmények. Hosszuk azonban az 1 centimétert is elérheti. Az egész izomban találhatók elszórva, és az izom hirtelen megnyújtásakor elektromos impulzus formájában információt küldenek a központi idegrendszer gerincvelői része felé.
Az izomorsót (4.10. ábra) kívülről az extrafuzális rostok (extrafusal fibers) veszik körbe. Ezek rögzítik az izomorsót az izom többi részéhez, így a nyújtásból adódó mechanikai erő (feszülés) közvetítésében játszanak szerepet. Az izomorsón belül a feszülésre érzékeny intrafuzális rostok találhatók. Ezeknek három típusa van: 1. dinamikus magzsákrost (dynamic nuclear bag), 2. statikus magzsákrost (static nuclear bag), 3. magláncrost (nuclear chain fiber). Mindhárom intrafuzális rost középső részét az Ia típusú afferens axon elágazó idegvégződései veszik körbe, melyek spriálszerűen rátekerednek a rostokra. A II-es típusú afferens axon azonban csak a statikus magzsákrostra és a magláncrostra tekeredik rá, tehát kihagyják a dinamikus magzsákrostot (ennek fontos szerepe van a nyújtás sebességének megkülönböztetésében). Ha a teljes izmot eredésénél és tapadásánál fogva megnyújtjuk, akkor az extrafuzális rostokkal együtt az intrafuzális rostok is megnyúlnak, és így az Ia és a II-es axonvégződésekben ingerület váltódik ki, mely akciós potenciálként elindul a gerincvelő hátulsó szarvába. Az ingerület egyetlen szinapszissal áttevődik a gerincvelő elülső szarvában található, úgynevezett α motoneuronra (vagy α efferens), amely a megnyújtott izomhoz visszaküldi az ingerületet és kontrakciót vált ki. A reflexkör, amely egy afferentációból és egy efferentációból áll, ezzel bezárul. A nyújtási reflex tehát az izom nyújtása miatt bekövetkező izomfeszülés, amely egyrészt az izom túlnyújtása elleni védelmet biztosítja, másrészt a testtartásban és az összetettebb mozgásoknál (pl. futás, ugrás) a koordinációban játszik fontos szerepet.
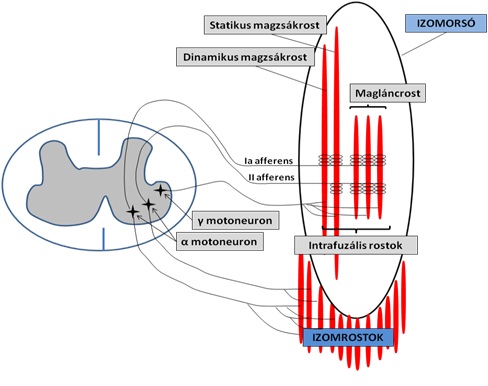
4.10. ábra. Az izomorsó és a nyújtási reflex működési sémája. Az izomorsó megnyújtásával az intrafuzális rostok (statikus magzsákrost, dinamikus magzsákrost, magláncrost) aktiválják az afferens axonokat (Ia és II-es axon). Az afferenseken haladó ingerület a gerincvelőben áttevődik az ? motoneuronokra, amelyek így magukat az izomrostokat aktiválják, az izom megfeszül. A α motoneuron az intrafuzális rostok érzékenységét állítja be, mely különböző edzésprogramok hatására jelentősen változhat.
A fentiekben említett reflexkör meg kell, hogy különböztesse a nagy sebességű (fázisos) nyújtást, valamint a nyújtás utáni állandósult, „megnyújtott” (statikus) állapotot. A nagy sebességű nyújtásokat mindhárom intrafuzális rost érzékeli, és az azokat körbevevő Ia afferensen elindulnak az akciós potenciálok a gerincvelőbe, majd az átkapcsolás után vissza az izomba. Minél nagyobb a nyújtás sebessége, annál nagyobb frekvenciával indulnak az akciós potenciálok, hiszen az izomnak annál nagyobb erővel kell majd ellenállnia a nyújtásnak. Tehát minél nagyobb sebességgel nyújtjuk az izmot, annál erősebb reflexválaszt kapunk (Kyrolainen és Komi, 1994). Ha azonban a megnyújtott állapotot megtartjuk, azt már csak a statikus magzsákrost érzékeli, amelyből lényegesen kisebb frekvenciájú akciós potenciálok indulnak ki a gerincvelőbe és onnan az izomba.
4.4.2. Az ínorsó működése: a Golgi reflex
Az ínorsók, vagy az ismert Camillo Golgi után elnevezett Golgi készülékek (Golgi tendon organ) olyan izomreceptorok, amelyek lényegesebben egyszerűbb szerkezettel és működési mechanizmussal rendelkeznek, mint az izomorsók. Maximális hosszuk mehközelítően 1 mm, átmérőjük 0,2 mm, tehát kisebbek, mint az izomorsók. Az ínorsók az izom és az ín átmeneti határán helyezkednek el. Valamennyi ínorsó kapcsolódik néhány izomrosthoz, ezek közvetítik az erőt az ínorsóra. Az ínorsót egyetlen axon, úgynevezett Ib axon hálózza be, és az ín feszülésekor ez közvetíti az információt a gerincvelőbe (4.11. ábra).
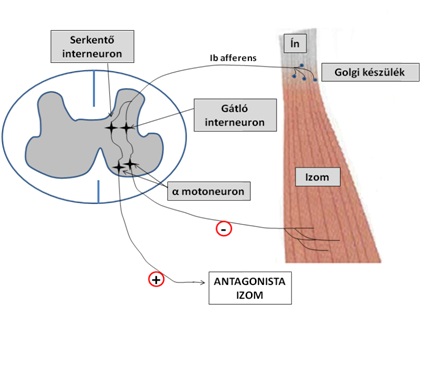
4.11. ábra. A Golgi reflex működési sémája. Az agonista izom aktív erőkifejtésekor a Golgi készülékben ingerület keletkezik, mely az Ib afferensen keresztül a gerincvelő hátulsó szarvba jut, és egy serkentő interneuronon keresztül az antagonista izmot aktiváló α motoneuronra tevődik át (+): az antagonista izom aktiválódik (antagonista co-kontrakció). Ugyanakkor az agonista izomhoz a gátló interneuronon keresztül nem jut további ingerület, megakadályozva az agonistát veszélyeztető további feszülést (-)
Az ínorsók működésére jellemző, hogy elsősorban nem az izom passzív nyújtására érzékenyek, hanem az izom akaratlagos megfeszítésekor bekövetkező, az inak megnyújtását előidéző megnyújtásra. Az ínorsók az izomrostokon keresztül mechanikai kapcsolatban állnak az egyes motoros egységekkel. Ha egy motoros egység aktív, akkor a hozzá tartozó ínorsó erről információt küld a központi idegrendszerbe. Ez a kapcsolat azt teszi lehetővé, hogy erőkifejtésünk mértékét szabályozni, kontrollálni tudjuk. Olyan mozgásoknál, vagy testtartásnál, ahol folyamatosan ugyanakkora erőt kell kifejtenünk (pl. karok megemelése és megtartása mozdulatlanul, vagy fekvőtámasz megtartása), az ínorsók által érzékelt feszülésből a központi idegrendszer „informálódik” (afferentáció) és szabályozza, hogy mennyi motoros egységet aktiváljon (efferentáció) a kívánt erőszint fenntartásához.
Az ínorsók másik jelentősége abban rejlik, hogy a teljes izom nagy erejű megfeszítésekor gátolja egyes motoros egységek működését, tehát megakadályozza az izom további feszítését. Ugyanakkor facilitálja (serkenti) az ellenoldali (antagonista) izom motoros egységeit, hogy az agonista és antagonista izom által érintett ízületet stabilizálja. Túl nagy agonista erőkifejtés esetén ugyanis az ízületben olyan elmozdulás történhet, ami károsítja az ízület alkotórészeit. Ezt a jelenséget, tehát amikor az agonista izom aktiválásával az antagonista izom is kismértékben aktiválódik antagonista co-kontrakciónak nevezzük (antagonist co-activation). Minél nagyobb erővel feszítjük meg az egyik izmot, az antagonista annál nagyobb erővel fog aktiválódni.
A Golgi-reflex működése nyújt segítséget a közismert, nyújtó hatású gimnasztikában alkalmazott PNF módszerben (posztizometriás neuromuszkuláris facilitáció), melyet gyakran alkalmaznak az edzők, terapeuták (Hindle és mtsai, 2012). A PNF három fázisból áll: először az érintett izmot néhány másodpercig statikusan teljes erővel megfeszítjük, majd ellazítjuk, végül pedig statikusan nyújtjuk. A statikus előfeszítésre azért van szükség, mert hatására az ínorsók érzékenysége átállítódik. Így az ellazítás után következő statikus nyújtáskor kisebb a reflextevékenység, tehát kedvezőbb a nyújtó hatás.
4.4.3. A nyújtási reflex szerepe a mozgásszabályozásban
A nyújtási reflex és a Golgi reflex mechanizmusának ismerete fontos ahhoz, hogy megértsük az egyes bemelegítő és nyújtó hatású gimnasztikai gyakorlatok fontosságát és különbözőségét. Ballisztikus, vagy utánmozgásos gimnasztikai gyakorlatoknál az izom nyújtása gyors és rövid ideig tart, ebben az esetben az izom erős kontrakcióval fog válaszolni. Ezért ezek a gyakorlatok kevésbé alkalmasak az ízületi mozgástartomány növeléséhez, inkább bemelegítésre alkalmazzák a sportolók. Statikus nyújtáskor (stretching) azonban csak a statikus magzsákrost vált ki reflexet, így az izom lényegesen kisebb erejű kontrakcióval fog ellenállni a nyújtásnak. Ráadásul, ha a statikusan nyújtott állapotot hosszabb ideig (akár 20-30 másodpercig) megtartjuk, a magzsákrostból érkező afferens impulzusok egyre kisebb frekvenciával fognak érkezni, ennek következtében értelemszerűen az izom jobban ellazul. Ezek a gyakorlatok így alkalmasabbak a mozgástartomány növeléséhez, rendszeres gyakorlással jelentősen jelentős javulás érhető el. A stretchinget kisebb mértékben alkalmazzák bemelegítésre, inkább különálló edzésrészként (általában az edzések végén) végzik a sportolók.
A nyújtási reflex kiváltható olyan gyakorlatokkal, amelyek nyújtásos-rövidüléses kontrakciókból állnak. Ezt a kontrakció típust már korábban jellemeztük, és ismertettük, hogy egy rövid ideig tartó és gyors excentrikus, majd egy azt követő koncentrikus fázisból áll. Ilyen gyakorlatok a plyometriás gyakorlatok, vagyis az összes reaktív erőt fejlesztő ugró- és szökdelőgyakorlat. Amennyiben az izom nyújtása (vagyis az excentrikus fázis) ezeknél a gyakorlatoknál elég gyors, akkor működésbe lép a nyújtási reflex. Ezt úgy érhetjük el, ha a súlypontunkat gyorsan süllyesztjük az ugrás excentrikus fázisában, lassú súlypontsüllyesztéssel ugyanis nem váltódik ki a reflex. Valamennyi nyújtásos-rövidüléses kontrakció során, ahol a nyújtás fázisa gyors, nyújtási reflex jön létre, és ez pozitívan befolyásolja az izom koncentrikus munkavégzését.
Instabil helyzetekben, amikor az egyensúlyvesztés lehetősége fenn áll, a nyújtási reflexnek fontos szerepe van a testtartás szabályozásában. Különböző statikus és dinamikus instabil támaszhelyzetekben az agonista és antagonista izmokban létrejövő erőkifejtés aránya határozza meg azt, hogy az instabil helyzetet fenn tudjuk-e tartani. Egy lábon állásnál például a frontális síkban a boka supinátor és pronátor izmai, míg a szagitális (oldal) síkban a plantarflexor és dorsalflexor izmok egyidejű működése stabilizálja a bokaízületet. A boka hirtelen billenésekor az egyik oldalon az izmok hirtelen megnyúlnak, ami kiváltja a nyújtási reflexet és az izom gyors aktivációját, ami stabilizálja a bokát. Dinamikus egyensúly megtartásában, mint például labdajátékokban végrehajtott irányváltásoknál, vagy egy lábról végzett ugrásoknál és leérkezéseknél is fontos szerepet tölt be a nyújtási reflexek bekapcsolódása. A közkedvelt proprioceptív tréninget (proprioceptive training) ennek a képességnek a fejlesztésére alkalmazzák. Ezek olyan statikus és dinamikus egyensúlygyakorlatok, amelyeket általában instabil felületeten, vagy instabil eszközök (pl. egyensúlyozó korong) segítségével végzik.
4.4.4. Az izomorsó érzékenységének beállítása
A nyújtási reflex mechanizmusában fontos szerepet játszik az úgynevezett γ axon (vagy gamma efferens). A γ axonok a gerincvelő szintén elülső szarvában található γ motoneuronokból indulnak ki, és az izomorsó intrafuzális rostjaihoz futnak (4.10. ábra). Az intrafuzális rostok érzékenységének beállítását végzik, tehát azt szabályozzák, hogy az izom mekkora sebességű és milyen erősségű nyújtásra reagáljon kontrakcióval. A γ efferens, az Ia/II afferens, valamint az α efferens pályát γ huroknak nevezzük.
A nyújtási reflex számos edzésprogram hatására módosulhat akut vagy krónikus módon. Az egyszer elvégzett edzések után (akut hatás) a reflexválasz azonban nem csupán a gamma hurok átállításával módosulhat, hanem különböző kémiai (pl. kalciumkiáramlás) és mechanikai (pl. mikrosérülések) okok miatt változik. A kutatók azt tapasztalták, hogy közvetlenül megerőltető állóképességi tevékenység (maratoni futás) után kisebb a reflexválasz, ami a neuromuszkuláris rendszer fáradásával magyarázható (Avela és mtsai 1999a). Ez az állapot néhány napig fennmaradhat, amely már viszont az izom mikrosérüléseinek köszönhető. Hasonlóan csökken az izom reflexválasza hosszantartó stretching után, melyet azzal magyaráznak a kutatók, hogy az izomorsóban található intrafuzális rostok nyúlékonyabbakká válnak (Avela és mtsai 1999b). A reflexválasz erőssége függ az edzettségi háttértől. 16 hétig tartó erőfejlesztő edzés után csökken a reflex ereje, valószínűleg az izomorsók érzékenységének csökkenése miatt (Hakkinen és Komi, 1983). Állóképességi sportolóknál nagyobb izomorsó érzékenységet tapasztaltak, mint erősportot végző személyeknél (Kyrolainen és Komi, 1994).
ÖSSZEFOGLALÁS
- Anyagcsere folyamatok alapján lassú oxidatív (SO), gyors oxidatív-glikolítikus (FOG) és gyors glikolítikus (FF) izomrostokat különböztetünk meg.
- Miozin ATP-áz aktivitás alapján hét izomrostot különítünk el: I, IC, IIC, IIAC, IIA, IIAB és IIB. Ebben a sorrendben ezeknek a rostoknak a kontraktilitási sebessége és fáradási hajlama nő.
- Motoros egységnek nevezzük egy ? motoneuron és az általa beidegzett izomrostok összességét. A motoros egységek egyidejű bekapcsolásával, melyet intramuszkuláris koordinációnak nevezünk, fokozhatjuk erőkifejtésünket, csökkenthetjük az erőkifejtés idejét.
- Fáradás szempontjából háromféle motoros egység típust különböztetünk meg: lassú (S), gyors fáradástűrő (FR) és gyors fáradékony (FF). A gyors motoros egységekhez vastagabb axon, vastagabb izomrost és több izomrost tartozik, így nagyobb és gyorsabb erőkifejtésre képesek a lassúakkal szemben.
- Ha erőkifejtésünk mértékét fokozatosan növeljük, akkor először a kisebb, lassabb egységek kapcsolódnak be, majd később a nagyobb, gyors egységek is. Ezt a jelenséget „méretelvnek” nevezzük.
- Az izmokhoz érkező elektromos impulzusok együttesét az izom elektromos aktivitásának nevezzük, melyet elektromiográfiával (EMG) vizsgálunk. Az EMG jel nagysága (amplitúdója) gyakran összefügg az izom erőkifejtésével.
- Az izom hirtelen megnyújtásakor az izom egyes feszülésérzékeny receptoraiból (izomorsókból) származó ingerület hatására az izom kontrakcióval válaszol, melyet nyújtási, vagy miotatikus reflexnek nevezünk.
- Az agonista izom nagyerejű megfeszítésekor a Golgi-reflex gátolja az izom további aktivációját, ugyanakkor serkenti az antagonista izom aktivációját, melyet antagonista co-kontrakciónak nevezünk.
Irodalomjegyzék
Aagaard P, Andersen JL (1998) Correlation between contractile strength and myosin heavy chain isoform composition in human skeletal muscle. Medicine and Science in Sports and Exercise, 30, 1217-1222.
Avela J, Kyröläinen H , Komi PV , Rama D (1999a) Reduced reflex sensitivity persists several days after long-lasting stretch-shortening cycle exercise. Journal of Applied Physiology, 86, 1292-1300.
Avela J, Kyröläinen H , Komi PV (1999b) Altered reflex sensitivity after repeated and prolonged passive muscle stretching. Journal of Applied Physiology, 86, 1283-1291.
Armstrong RB, Saubert CW, Seeherman HJ, Taylor CR (1982) Distribution of fiber types in locomotory muscles of dog. American journal of anatomy, 163, 87-98.
Bodine SC, Roy RR, Elder E, Edgerton VR (1987) Maximal force as a function of anatomical features of motor unit sin the cat tibialis anterior. Journal of Neurophysiology, 6, 1730-1745.
Brooke MH, Kaiser KK (1970) Muscle fiber types: How many and what kind? Archives of Neurology, 23, 368-379.
Burke RE (1967) Motor unit types of cat triceps surae muscle. Journal of Physiology (London), 193, 141-160.
Engel WK (1962) The essentiality of histo- and cytochemical studies of skeletal muscle int he investigation of neuromuscular disease. Neurology, 12, 778-784.
Gollnick PD, Sjödin B, Karlsson J, Jansson E, Saltin B (1974) Human soleus muscle: A comparison of fiber composition and enzyme activities with other leg muscles. Pflügers Archiv, 348, 247-255.
Hakkinen K, Komi PV (1983) Changes in neuromuscular performance in voluntary and reflex contraction during strength training in man. International Journal of Sports Medicine, 4, 282-288.
Henneman E, Somjen G, Carpenter DO (1965) Functional significance of cell size of spinal motoneurons. Journal of Neurophysiology, 28, 560-580.
Hindle KB, Whitcomb, Briggs WO, Hong J (2012) Proprioceptive Neuromuscular Facilitation (PNF): Its Mechanisms and Effects on Range of Motion and Muscular Function. Journal of Human Kinetics, 31, 105-113.
Johnson MA, Polgar J, Weightman D, Appleton D (1973) Data on the distribution of fiber types in thirty-six human muscles. Journal of the Neurological Sciences, 18, 111-129.
Kyrolainen H, Komi PV (1994) Stretch feflex responses following mechanical stimulation in power and endurance trained athletes. International Journal of Sport Medicine. 15, 290-294.
Lieber RL (2002) Skeletal muscle structure, function, and plasticity. 2. kiadás, Lippincott Williams and Wilkins, Baltimore, Philadelphia, pp 86-87.
Lowey S, Waller GS, Trybus KM (1993) Skeletal muscle myosin light chains are essential for physiological speeds of shortening. Nature, 365, 454-456.
Martin TP, Bodine-Flower S, Roy RR, Eldred E, Edgerton VR (1988) Metabolic and fiber size properties of cat tibialis anterior motor units. American Journal of Physiology, 255, C43-C50.
Peter JB, Barnard RJ, Edgerton VR, Gillespie CA, Stempel KE (1972) Metabolic profiles on three fiber types of skeletal muscle in guinea pigs and rabbits. Biochemistry, 11, 2627-2733.
Ranvier L (1873). Popriétés et structures différents des muscles rouges et des muscles blancs, chez les lapins et chez les raies. Comptes Rendus Hebdomadaires des Seances de l’Academie des Sciences: D. Sciences Naturelles (Paris), 77, 1030-1034.
Schiaffino S, Gorza L, Sartore S, Saggin L, Vianello M, Gundersen K, Lomo T (1989) Three myosin heavy chain isoforms in type 2 skeletal muscle fibers. Journal of Muscle Research and Cell Motility, 10, 197-205.
Staron RS, Hikida RS (1992) Histochemical, biochemical, and ultrastructural analyses of single human muscle fibers, with special reference to the C-fiber population. The Journal of Histochemistry and Cytochemistry, 40, 563-568.
VanBuren P, Waller GS, Harris DE, Trybus KM, Warshaw DM, Lowey S (1994) The essential light chain is required for full force production by skeletal muscle myosin. Proclamation of the National Academy of Science, 91, 12403-12407.
Zappe HA, Maeda Y (1985). X-ray diffraction study of fast and slow mammalian skeletal muscle in the live relaxed state. Journal of Molecular Biology, 185, 211-214.
| « Előző fejezet | Tartalomjegyzék | Következő fejezet » |
Események
Jelenleg nincs aktuális esemény.