TÁMOP-4.1.2.E-13/1/KONV-2013-0012
"Sporttudományi képzés fejlesztése
a Dunántúlon"
- -> Tananyagfejlesztés
- -> A vázizom...
- -> 3.












3.
3. A VÁZIZOM MŰKÖDÉSÉNEK ÉLETTANA
- 3.1 Az ingerlés – kontrakció folyamata
- 3.1.1. Az ingerület érkezése a motoneuronon
- 3.1.2. Az ingerület átterjedése az izomrost membránjára
- 3.1.3. Az ingerület bejutása az izomrost belsejébe
- 3.1.4. A Ca+ kiáramlása a szarkoplazmatikus retikulumból
- 3.1.5. Az aktomiozin kötés kialakulása
- 3.1.6. A kereszthídciklus
- 3.1.7. A Ca+ visszaáramlása a szarkoplazmatikus retikulumba
- 3.2. Izomrángás, tetanusz, elektromechanikai késés
- 3.3. Kontrakció típusok
A fentiekben részletesen olvashattunk a harántcsíkolt izom szerkezetéről, és a teljes izom alakjától kiindulva a molekuláris szerkezetig tanulmányozva világossá vált, hogy az izom erőkifejtésért felelős szövet. A következőkben betekintést nyerhetünk abba a lenyűgöző molekuláris mechanizmusba, amely során az izomrostok a tápanyagokból származó kémiai energia felhasználásával mechanikai munkát képesek végezni.
3.1 Az ingerlés – kontrakció folyamata
Az ingerlés – kontrakció folyamat (excitation – contraction coupling) alatt azt a mechanizmust értjük, amely során a központi idegrendszerből érkező elektromos impulzus (ingerület) hatására bekövetkezik az izomkontrakció. Ez a folyamat több fázisra osztható, melyeket az alábbiakban ismertetünk.
3.1.1. Az ingerület érkezése a motoneuronon
Ahogyan azt már korábban említettük, az izmok működéséhez idegrendszeri vezérlésre van szükség. Fiziológiás körülmények között valamennyi izomrostot beidegez egy mozgató idegsejt (motoneuron), és ennek az idegsejtnek a nyúlványán (axon) érkező akciós potenciál fogja elindítani az izomkontrakciót. A felső motoneuronok az agykéregben, az alsó motoneouronok pedig a nyúltvelőben és a gerincvelőben találhatók. A felsők a motoros kéregből továbbítják az információt az izmokhoz az alsó motoros idegeken keresztül. Az akciós potenciálok (action potentials) olyan elektromos impulzusok, melyek nagyon nagy sebességgel haladnak az axonokon. Az axonon a sejtmembrán két oldalán található ionok (töltéssel rendelkező részecskék) hirtelen átáramlása megváltoztatja a membránfeszültség polaritását (depolarizáció), és ez hullámszerűen terjed végig.
Maga a kontrakció elindítása történhet akaratlagosan, vagy reflexmechanizmus által. Akaratlagos kontrakció esetén az elektromos ingerület az agykéregből indul ki és több idegsejten keresztül (átkapcsolások mentén) jut el a célzott izomrostokig. Reflex által kiváltott kontrakciós esetében az ingerület kiindulópontja a gerincvelő, és innen az ingerület átkapcsolás nélkül jut el a célrostokig. Fiziológiás körülmények között beidegzés nélkül az izmok nem működtethetők, azok csak külső elektrostimuláció segítségével ingerelhetők. Az izom bénulása általában akkor következik be, ha az idegi összeköttetés megszűnik a központi idegrendszerrel. Amennyiben ez az állapot hosszú idegig fennmarad a denervált izom inaktivitás miatt sorvadásnak indul.
3.1.2. Az ingerület átterjedése az izomrost membránjára
Amikor az elektromos impulzus eléri az izomrost felszínét, a neuromuszkuláris szinapszison (neuromuscular junction) keresztül áttevődik a rost membránjára, a szarkolemmára. A neuromuszkuláris szinapszist az axonvég membránja, az izomsejt membránja, valamint a kettő közötti szinaptikus rés alkotja. Az izomsejthez tartozó membránterületet, amelyhez az axon kapcsolódik motoros véglemeznek (motor endplate) nevezzük. Az axon az izomrosthoz érve szerteágazik, úgynevezett terminális ágacskákat létrehozva. Ezekben a részben találhatók a szinaptikus hólyagocskák (synaptic vescicles), melyek acetil-kolin neurotranszmittert (ingerületátvivő anyag) tartalmaznak. Az elektromos impulzus érkezésekor a hólyagocskák kiszabadulnak az axonból és a szinaptikus résbe (synaptic cleft) áramlanak. Ez a folyamat megnyitja a motoros véglemez Na+ csatornáit, ami az izommembránon elindítja a tovaterjedő akciós potenciált. Fiziológiás esetben egy motoros ideg akciós potenciál egy izomrost akciós potenciált vált ki, de különböző kóros állapotoknál ez megváltozhat.
3.1.3. Az ingerület bejutása az izomrost belsejébe
Az izomrost membránján immáron végighaladó ingerület valamilyen módon be kell, hogy jusson a rost belsejébe. Erre szolgálnak a T-tubulusok, amelyek a membrán izomrostra merőleges, szabályosan ismétlődő betüremkedései (Peachey 1965). A T-tubulusok a szarkomerek A és I csíkjának találkozásánál türemkednek be, így minden szarkomerhez két T-tubulus tartozik. A T-tubulusok nemcsak betüremkednek, hanem körbe is veszik az egyes myofibrillumokat, biztosítva azt, hogy az ingerület valamennyi kontraktilis fehérjéhez eljusson. Így tulajdonképpen az egész izomrostot behálózzák.
3.1.4. A Ca+ kiáramlása a szarkoplazmatikus retikulumból
A T-tubulusokon végighaladó akciós potenciál a T-tubulusok közvetlen szomszédságában elhelyezkedő szarkoplazmatikus retikulumból Ca+ kiáramlást kezdeményez, amely a kontrakció elindító kulcsa. A Ca+ ion kontrakcióban betöltött szerepét már több, mint száz éve felismerték a kutatók (Ringer 1883, Mines 1913). De mégis hogyan tevődik át az ingerület a T-tubulusokból a szarkoplazmatikus retikulumokra? A T-tubulosokban találhatók az úgynevezett dihidropiridin (DHPR) receptorok, melyek érzékenyek a feszültségváltozásra. Ezekben a receptorokban az akciós potenciál érkezésekor konformációváltozás (szerkezeti és alaki változás a molekulákban) következik be. Ezt a konformációváltozást érzékelik a közvetlen szomszédságban lévő szarkoplazmatikus retikulumok rianodin receptorai, melyek Ca+ csarornái ekkor megnyílnak, kiáramoltatva a Ca+ ionokat az izomrost plazmájába (szarkoplazma). A Ca+ ionok kiáramlása passzív, tehát energia igény nélküli folyamat.
3.1.5. Az aktomiozin kötés kialakulása
A szarkoplazmatikus retikulumból szabadon kiáramló Ca+ ionok immáron könnyedén elérik a kontraktilis fehérjéket. Amikor az izom nyugalomban van, vagyis amikor a plazma Ca+ koncentrációja alacsony, a tropomiozin fehérje az aktin teljes hosszában elfoglalja annak miozinkötő helyeit, tehát nem jöhet létre kereszthíd (az előző fejezetben már tárgyaltuk, hogy a tropomiozin az aktinon végighúzódó, kontrakciót szabályozó fehérje). A Ca+ kiáramlásakor azonban a Ca+ a tropomiozinon lévő troponin C-hez kötődik, melynek hatására a tropomiozin elfordul olyannyira, hogy az aktinon található miozinkötő helyek felszabadulnak. Ebben a pillanatban a miozin feje az aktinhoz kötődik létrehozva a kereszthidat és a kontrakciót. Mindaddig, amíg Ca+ jelen van, ez a folyamat végbemegy. Amennyiben nem érkeznek akciós potenciálok, a Ca+ visszakerül a szarkoplazmatikus retikulumba. Ez a folyamat aktív, tehát ATP-t igénylő transzport folyamat, és a SERCA (szarkoplazmatikus/endoplazmatikus retikulum Ca-ATP-áz) nevű pumpa működteti.
3.1.6. A kereszthídciklus
Eljutottunk tehát a kereszthíd kialakulásáig, vagyis hogy egy elektromos jel hatására létrejöjjön az aktomiozin kötés. Az izomműködés talán egyik legérdekesebb folyamata az, hogy mégis hogyan jön létre mechanikai munkavégzés? A csúszófilamentum mechanizmusnál már említettük, hogy a mechanikai munka az aktin és miozin filamentumok egymással ellentétes irányba történő elcsúszása révén jön létre, mely elcsúszást a kereszthidakban kialakuló konformációváltozás (molekuláris szerkezetváltozás) hozza létre. Ennek folyamatát számos kutató tanulmányozta, és több elmélet is született. A csúszófilamentum mechanizmus felfedezését követően A. F. Huxley (1974) később arra is rájött, hogy az aktin és miozin fehérjék folyamatos elcsúszását a kereszthidak ciklikus létrejötte és felbomlása hozza létre. Lymn és Taylor (1971) szerint az aktin és miozin közötti kötés lehet „erős” és „gyenge”. A. F. Huxley és Simmons (1971) pedig igazolták, hogy maga az elcsúszás a miozin fej elfordulásának tulajdonítható. Ezeket az elméleteket Stein és mtsai (1979) elméleteivel kiegészítve a kereszthídciklus az alábbiak szerint modellezhető (3.1. ábra):
1. Az aktin-miozin közötti erős kötést (strong binding state) „rigor” (merev) állapotnak nevezzük, és a ciklus elemzését általában ettől a ponttól szokták kezdeni (3.1. ábra). A miozin feje ilyenkor 45°-os szöget zár be a miozin rúdjával. A vázizom szerkezetének bemutatásakor már értelmeztük az erős kötést: erős akto-miozin kötés akkor létesül, amikor a miozin fej legnagyobb molekulasúlyú (aktinkötő) szegmensének hasadéka teljesen zárva van.
2. Az ATP belép a miozinfej hasadékába, de nem teljesen, mert az adenin rész kívül marad. Ez azonban már elegendő ahhoz, hogy a miozinfej két szegmense kissé eltávolodjon egymástól, meggyengítve az aktinnal való kötést (weak binding state) (3.1. ábra).
3. Az ATP-t teljesen egészében bejut a miozinfej hasadékába. Ekkor a miozinfej leválik az aktinról és a rúd és a fej közötti szög 45°-ról 90°-ra változik, ami kb. 5 nm elmozdulást jelent az aktin mentén: a miozinfej az aktin egy új kötőhelyével néz szembe (3.1. ábra). Az ATP ez alatt már ADP-re és szervetlen foszfátra (Pi) bomlott szét az ATP-áz enzim segítségével.
4. A szervetlen foszfát leválik az ATP-áz enzimről és így a miozinfej ismét kapcsolódhat az aktinhoz. Létrejön az erőcsapás (power stroke), vagyis a miozinfej és a rúd közötti szög ismét 45°-os lesz (3.1. ábra). Az erőcsapás alatt leválik az ADP is.
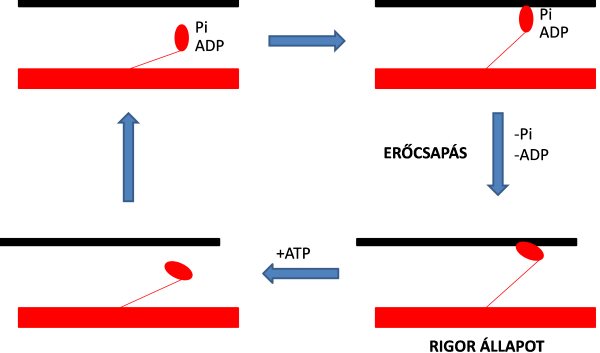
3.1. ábra. A kereszthídciklus folyamata (négyfázisos modell).
Egy kereszthíd ciklusban létrejött erőcsapás elmozdulása kb. 5 nm, erőkifejtése pedig kb. 5-10 pN (lásd az összefoglalót: Cook 1997).
3.1.7. A Ca+ visszaáramlása a szarkoplazmatikus retikulumba
A kontrakció mindaddig végbemegy, amíg Ca+ van jelen. Az elektromos impulzusok megszűnésekor a Ca+ visszakerül a szarkoplazmatikus retikulumba (Ca+ pumpa), mely aktív, tehát ATP függő transzporttal valósul meg. A gyors izomrostokban, a Ca+ pumpa rendkívül hatékony, ami gyors relaxációt jelent.
3.2. Izomrángás, tetanusz, elektromechanikai késés
A fentiekben szemléltettük, hogy egyetlen akciós potenciál érkezésekor az izomban hogyan jön létre a kereszthíd ciklus (kontrakció) és hogyan szűnik az meg (relaxáció). Egyetlen idegi impulzus nem elegendő tartós izom-összehúzódáshoz és terhek megemeléséhez. Ha az izomhoz egyetlen impulzus érkezne, csupán egy izomrángást tapasztalnánk (3.2. ábra). Izomrángás (muscle twitch) alatt egy idegi impulzus által létrehozott izomerő-kifejtést értjük. Akaratlagos kontrakcióknál egyetlen impulzus és az arra bekövetkező izomrángás kiváltása lehetetlen, azt csak külső elektrostimulációs berendezéssel tudjuk létrehozni. A fentiekben említettük, hogy egy idegi impulzus hatására bekövetkező kontrakció és relaxáció tehát ~100ms-ig tart. Mi történik azonban, ha ezen az időintervallumon belül, vagyis a teljes relaxáció előtt egy következő impulzus is érkezik a myofibrillumokhoz? A két impulzus felerősíti egymást, vagyis az izom nagyobb erőt fog kifejteni. Ennek okát az izom mechanikai működésében kell keresnünk: Az első impulzus érkezése következtében a kereszthidakban létrejön az erőcsapás és a szarkomerekkel sorba kapcsolt elasztikus fehérjék megnyúlnak. Az erőcsapás révén bár az aktin és miozin molekulák elmozdulnak egymás mentén, a Z vonalak igazából csak minimálisan közelednek egymáshoz a soros elasztikus elemek megnyúlása miatt. Még a relaxáció előtt érkező második impulzus azonban már valós rövidülést okoz a szarkomerekben, növelve az erőkifejtés mértékét. Még a relaxációs időn belül érkező, két egymás után következő impulzus együttes hatását szummációnak (temporal summation) nevezzük. Relaxációs időn belül érkező több egymás utáni impulzus még tovább növeli az izom erőkifejtését. Ha a sorozatimpulzusok érkezése alacsony frekvenciájú (pl. 10Hz), akkor az erőkifejtés görbén még láthatók az egyéni izomrángásból származó tüskék (3.2. ábra). Ezt inkomplett tetanuszos összehúzódásnak (unfused tetanic contraction) nevezzük. Ha azonban az impulzusok 100Hz-nél sűrűbben érkeznek, akkor olyan nagymértékű lesz a kalcium kiáramlás, hogy a szarkomerekben nem jön létre relaxáció és komplett tetanuszos összehúzódás következik (fused tetanic contraction) be (3.2. ábra). Ilyenkor az erőkifejtés folyamatos lesz.
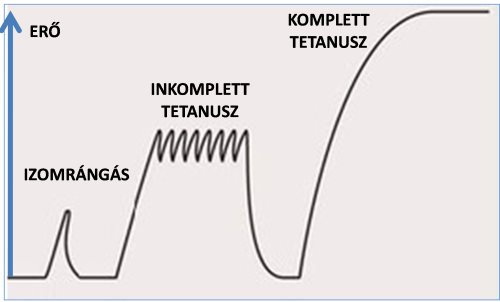
3.2. ábra. Az izom erőkifejtése egy (izomrángás), több alacsonyfrekvenciájú (inkomplett tetanusz) és több magas frekvenciájú (komplett tetanusz) impulzus érkezésekor.
Az erőkifejtés mértéke az elektromos impulzusok frekvenciájától függ, melyet kisülési, vagy tüzelési frekvenciának (firing rate) is nevezünk. A sorozatimpulzusok minél nagyobb frekvenciával érkeznek, annál nagyobb erőt fejt ki az izom, hiszen minél több kalcium áramlik ki a szarkomerekben annál több kereszthíd létesül. A tetanuszos izomkontrakciónak nagy jelentősége van a mozgás szabályozásában. A központi idegrendszer az impulzusok frekvenciájának variálásával az erőkifejtés mértékének változtatását teszi lehetővé. Ezt frekvenciakódolásnak nevezzük (rate coding). A célfeladattól függően erősebb vagy gyengébb erőkifejtésre van szükségünk, melyet a frekvencia növelésével vagy csökkentésével szabályozunk. Amennyiben a célfeladat elvégzésekor túl nagy vagy túl kicsi erőt fejtenek ki az izmok, a végrehajtás pontatlan, vagy nem gazdaságos. A finom és durva mozgásokat tartalmazó sportágaknál ennek kiemelkedő jelentősége van. Asztaliteniszben például teljes erejű leütéseket és finomabb nyeséseket is végre kell hajtani, mindezt a meglehetősen kis tömegű labdával és az asztalra pontosan helyezve. Az erőkifejtés ilyen magas szintű szabályozása tanulással, gyakorlással sajátítható el.
A fentiekben látható, hogy magát az ingerlést és kontrakciót alkotó biokémiai és mechanikai folyamatok időigényesek. Azt az időintervallumot, amely a motoneuronokból származó idegi impulzus elindulásakor kezdődik és az izom által az adott ízületben létrehozott mozgás megkezdéséig tart, elektromechanikai késésnek nevezzük (electromechanical delay) (Cavanagh és Komi, 1979). Az elektromechanikai késés függ:
- a sorbakapcsolt elasztikus elemek rugalmasságától (stiffness).
- az izom térfogatától
- az izom maximális erőkifejtő képességétől
- az izmot alkotó gyors és lassú rostok arányától
- az ingerületvezetés sebességétől.
Az elektromechanikai késés jelentősen nőhet fáradás következtében, hiszen ilyenkor mind a központi idegrendszerben, mind pedig az izomban biokémiai változások következnek be, amelyek miatt például lecsökken az ingerületvezetés és ingerület átvitel sebessége, vagy akár csökken a Ca pumpa hatékonysága. Mikrosérülések esetén is nőhet az elektromechanikai késés időtartama, hiszen egyes szarkomerek sérülnek és ideiglenesen kikapcsolnak, nem vesznek részt az erőkifejtésben. Az elektromechanikai késés jelentősen befolyásolja a reflexidőt.
3.3. Kontrakció típusok
Az emberi mozgásoknak számtalan variációi léteznek, melyek alatt a vázizom különböző módon képes erőt kifejteni, ezért különböző kontrakciótípusokat különítünk el. A kontrakciókat többféleképpen osztályozzák, például a szerint, hogy az izom milyen hosszváltozáson megy keresztül, miközben erőt fejt ki, vagy hogy mekkora gyorsulással és feszüléssel rendelkezik kontrakció alatt. Az egyes kontrakciótípusok definiálásához elsősorban Klavora (2007) művét használtuk fel.
3.3.1. Kontrakció típusok az izom hosszváltozásának tekintetében
Ha egy izom aktív, akkor annak hosszváltozását a külső ellenállás nagysága határozza meg. Amennyiben a külső ellenállás nagysága megegyezik az izom által kifejtett erővel, akkor izometriás, vagy statikus kontrakcióról (isometric contraction) beszélünk (a görög eredetű szó jelentése szerint izo = azonos, meter = hosszúság). Ilyenkor az izom úgy fejt ki erőt, hogy annak hossza, vagyis az eredése és tapadása közötti távolság nem változik. Ebben az esetben a csontok nem mozdulnak el egymáshoz képest, nem jön létre mozgás. Izometriás kontrakciók elsősorban a testtartásnál jelentkeznek pl. állás, vagy ülés közben. Izometriás erőkifejtést végzünk akkor is például, ha egy kézisúlyzót oldalsó középtartásban mozdulatlanul megtartunk. A sportmozgások közül elsősorban a torna statikus elemeinél (pl. mérlegállás, kézállás, ülőtámasz, függések) fejtenek ki az izmok erőt izometriás módon. Kötöttfogású birkózók mérkőzése során nagyon sok olyan szituáció van, amikor a versenyzők nem mozdulnak, ám a hosszú ideig tartó statikus erőkifejtések során rengeteg energiát elhasználnak.
Fontos megértenünk, hogy egy adott ellenállás ellenében kifejtett izometriás erőkifejtés közben a kontraktilis elemek, vagyis az aktív izomrészek valójában rövidülnek: aktin és miozin filamentumok elcsúsznak egymáson. A kontraktilis elemek által létrejött rövidülés mellett azonban az izom sorbakapcsolt elasztikus elemei megnyúlnak. Így a teher/ellenállás, melyre erőhatást gyakorolunk, nem mozdul el (3.3. ábra).
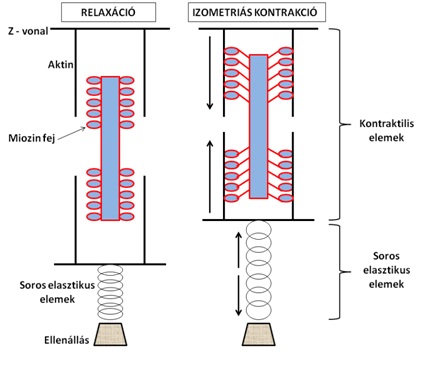
3.3. ábra. Kontraktilis és elasztikus izomrészek modellezése izometriás kontrakció közben. Az ábra bal oldalán található izom inaktív, aktin és miozin kereszthíd kapcsolat nem létesül. A jobb oldali izomnál az aktin és miozin filamentumok elcsúszása a kontraktilis apparátusban rövidülést okoz, de a soros elasztikus elemek megnyúlása miatt az ellenállást nem győzi le az izom (izometriás kontrakció).
Amikor az aktív izom változtatja hosszát, anizometriás, vagy dinamikus kontrakcióról (dynamic contraction) beszélünk. Dinamikusan az izom kétféle módon fejthet ki erőt: koncentrikusan és excentrikusan. Koncentirkus kontrakció (concentric contraction) akkor következik be, ha az erőkifejtés közben az izom rövidül, vagyis az eredése és a tapadása közötti távolság csökken. Ilyenkor az izom által kifejtett erő nagyobb a legyőzendő tehernél. Az izom eredésénél és tapadásánál fogva elforgatja az ízesülő csontokat az ízületi tengely(ek) körül, „legyőző” mozgás megy végbe. Súlyok megemelése (pl. kinyomása melltől fej fölé) vagy saját test megemelése (pl. guggolásból felállás, fekvőtámaszban a karnyújtás fázisa) közben az izmok koncentrikus erőt fejtenek ki. A 3.4 ábrán a könyökízületet áthidaló kétfejű karizom koncentrikus kontrakció alatt a felkart és alkart közelíti egymáshoz, így a könyökízület szöge csökken, a kézbentartott súlyt a könyökhajlító izom elmozdítja.
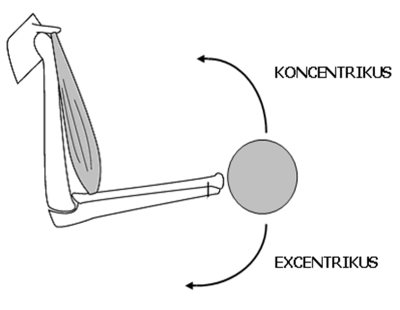
3.4. ábra. A kétfejű karizom rövidülése, vagyis eredési és tapadási pontjának közeledése (koncentrikus kontrakció) a súly megemelését eredményezi. Az izom nyúlása, vagyis eredési és tapadási pontjának távolodása (excentrikus kontrakció) pedig a súly leengedését eredményezi. A súly lassú, kontrolált leengedését az teszi lehetővé, hogy az izom ebben a fázisban is aktivált állapotban van, így a súly nem csupán a gravitációból származó erő hatására mozog.
A koncentrikus kontrakciót szemléltető izommodellt a 3.5. ábra mutatja be. Koncentrikus kontrakció elindításakor valójában az izom kontraktilis elemeiben rövidülés következik be, de az inak nyúlásával az ellenállás még nem mozdul el. Ez a pillanat egy izometriás előfeszülést jelent, hasonlóan az izometriás kontrakció izommodelljéhez. Mivel azonban az ellenállás kisebb, mint az izom ereje, ezért a kontraktilis elemekben bekövetkező további rövidülés miatt a teher legyőzésre kerül.
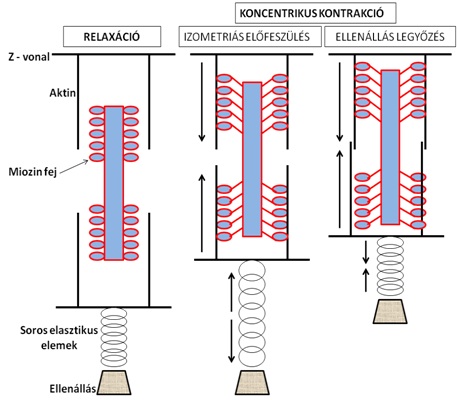
3.5. ábra. Kontraktilis és elasztikus izomrészek modellezése koncentrikus kontrakció közben. Az ábra bal oldalán található izom inaktív, aktin és miozin kereszthíd kapcsolat nem létesül. Az ábra középső részén az aktin és miozin filamentumok elcsúszása a kontraktilis apparátusban rövidülést okoz, de a soros elasztikus elemek megnyúlása miatt az ellenállást nem győzi le az izom (átmeneti izometriás előfeszülés). Az ábra jobb oldalán a kontraktilis apparátus további rövidülése az ellenállás legyőzését teszi lehetővé.
Excentrikus kontrakcióról (eccentric contraction) akkor beszélünk, amikor az izom erőkifejtés közben nyúlik, vagyis az eredése és a tapadása közötti távolság nő. Ekkor az izom által kifejtett erő kisebb, mint a legyőzendő teher, úgynevezett „fékező” mozgás következik be. Az excentrikus erőkifejtéssel végezzük a terhek leengedését, vagy pl. saját testünk súlypontjának süllyesztését. A 3.4. ábrán a könyökízületet áthidaló kétfejű karizom excentrikus kontrakciója a felkar és az alkar csontjainak távolodásával jár. A könyökízület szöge ekkor nő, a terhet leengedjük. Felhívjuk a figyelmet, hogy excentrikus kontrakció alatt az izom mindvégig aktív, elektromos impulzusok érik. Ez a terhek kontrollált leengedését teszi lehetővé. Amennyiben a terhet hirtelen leengedjük, „ledobjuk”, az izom teljesen ellazított (inaktív) állapotban van. Olyan helyzet is előfordul, hogy a teher súlya jóval nagyobb, mint amekkora súlyt az izomerő kifejtésével meg tudnánk tartani. Ebben az esetben maximális erőkifejtés mellett is csak leengedni vagyunk képesek a terhet, szintén excentrikus kontrakcióról beszélünk.
Az excentrikus kontrakció alatt mind a kontraktilis mind pedig a soros elasztikus izomelemek megnyúlnak (3.6. ábra). Kereszthidak ennek ellenére létesülnek, azonban azok nagy része a túl nagy feszülés miatt bomlik szét, és nem megy át a normál ATP függő szétkapcsolódáson. A tudományban egyetértés van azzal kapcsolatban, hogy a szokatlan edzés következtében kialakuló mikrosérülések és az azt követő izomfájdalom ennek a folyamatnak köszönhető. Erről azonban egy későbbi fejezetben írunk részletesebben.
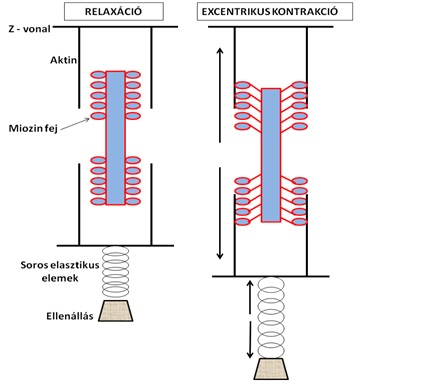
3.6. ábra. Kontraktilis és elasztikus izomrészek modellezése excentrikus kontrakció közben. Az ábra bal oldalán található izom inaktív, aktin és miozin kereszthíd kapcsolat nem létesül. A jobb oldali izommodellben bár kereszthidak létesülnek, az aktin és miozin filamentumok eltávolodnak egymástól, ami a kontraktilis apparátusban nyúlást okoz. Az ellenállás kontrollált elengedése következik be.
Korábban már említettük, hogy maga a latin eredetű kontrakció szó összehúzódást jelent, így megkérdőjelezhető, hogy izometriás és excentrikus működéskor miért használjuk a kontrakció szót. A biomechanika tudományban azonban a megértés megkönnyítéséért általánosan elfogadták a kontrakció terminus használatát minden olyan izomműködési módozatra, amelynél az izom aktív állapotban van. Az egyes kontrakció típusokat a hosszváltozás tekintetében a 3.7. ábra foglalja össze.
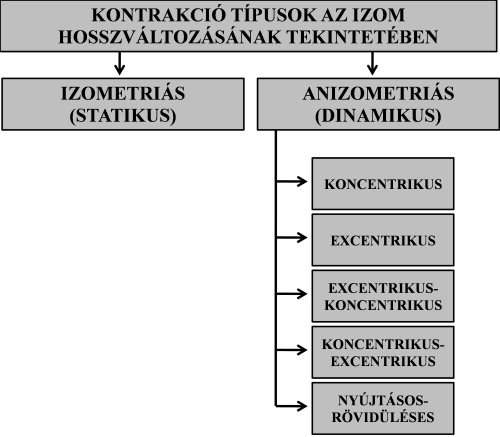
3.7. ábra. Kontrakció típusok az izom hosszváltozásának tekintetében.
3.3.2. Kontrakció típusok az izom feszülése és rövidülési/nyúlási sebessége tekintetében
Ennél a kategorizálásnál a kontrakciók alatt két tényezőt veszünk figyelembe: (1) kontrakció alatt hogyan változik a rövidülés vagy nyúlás sebessége, illetve (2) hogyan változik ez alatt az izom feszülése. A megértés megkönnyítése érdekében tisztázzuk, hogy az izom által kifejtett erő (force) és az izom feszülése (tension) alatt ugyanazt értjük, és a két fogalmat e szerint használjuk az egész tankönyvben.
Dinamikus kontrakcióknál az izom egy adott sebességgel rövidülhet, vagy nyúlhat. Amennyiben a rövidülés/nyúlás sebessége egy kontrakció során állandó, izokinetikus kontrakcióról (isokinetic contraction) beszélünk . Izokinetikus kontrakció természetes körülmények között nehezen hozható létre az emberi szervezetben. Ha pl. guggolásból megpróbálunk állásból állandó sebességgel állásba emelkedni, akkor az izmok megközelítőleg izokinetikus kontrakciót végeznek. Izokinetikus kontrakció inkább csak számítógép vezérlésű laboratóriumi erőmérő eszközökkel (dinamométer) szimulálható. A dinamométerek az erőfejlesztő gépekhez hasonló berendezések, viszont beprogramozhatók úgy, hogy a karok hajtásakor bármekkora erőt is fejt ki az egyén, a karok kizárólagosan az előre beállított szögsebességgel mozgathatók. Az ilyen kontrakciókat elsősorban tudományos kutatásoknál alkalmazzák, de a piacon már léteznek fitnesztermek számára készített verziók is. A dinamométerek működéséről a tankönyv utolsó fejezetben részletesen írunk. Az izokinetikus kontrakció egyébként koncentrikus és excentrikus kontrakció mellett is értelmezhető (pl. állásból ereszkedés állandó sebességgel guggolásba: izokinetikus excentrikus kontrakció). Izokinetikus kontrakciónál tehát az izom rövidülési/nyúlási sebessége nem változik, viszont az izom feszülése (erőkifejtése) pillanatról-pillanatra változik az ízületi szög és az izom hosszának változásával (magyarázat a későbbi fejezetekben).
Amikor az izom rövidülésének gyorsulása állandó, izotóniás kontrakcióról (isotonic contraction) beszélünk. Képzeljük el, hogy egy kipreparált izom egyik végét felfüggesztjük, másik végére pedig súlyt helyezünk. Ha az izmot folyamatosan ugyanakkora elektromos árammal stimuláljuk, az izom úgy fog rövidülni, hogy közben a legyőzendő teher gyorsulni fog. A teher folyamatos (elméletileg állandó) gyorsulása egészen egyszerűen Newton második törvényével magyarázható: F erő egy m tömegű testben a gyorsulást okoz (F = m · a). Modellünkben az azonos nagyságú elektromos stimuláció az izomban állandó erőkifejtést hoz létre (ha eltekintünk az izom hosszának változásától), és ez az erő az izom végére rögzített súlyt gyorsítja. Az izotóniás kontrakció olyan kontrakció, amelynél az izomban létrejövő feszülés is állandó (innen az elnevezés: izotónia = azonos feszülés). Az izotóniás kontrakció már nagymértékben hasonlít a természetes mozgásokhoz, de pontosan csak dinamométerrel tudjuk szimulálni, hasonlóan az izokinetikus kontrakcióhoz.
Az állandó gyorsulás tehát csak elméletileg létezik, a valós mozgatórendszerben csak megközelítőleg állandó a gyorsulás, ugyanis különböző szarkomerhossz mellett eltérő erőt képes az izom kifejteni, még ha ugyanakkora elektromos impulzus is éri. Továbbá az emberi szervezetben található emelőrendszereknek köszönhetően különböző ízületi szöghelyzetekben más és más forgatónyomatékot képesek az izmok kifejteni. Ezeknek mechanizmusát a későbbiekben tárgyaljuk. Az emberek és állatok mozgatórendszerére jellemző és természetesen előforduló kontrakció az auxotóniás kontrakció.Auxotóniás kontrakció (auxotonic contraction) alatt az izom rövidülési sebessége is és az izom feszülése is folyamatosan változik, éppen azért, mert ízületi szöghelyzettől függően változik az izom erőkifejtési képessége és rövidülési sebessége (megjegyzés: bár az auxotonia szó jelentése növekvő feszülés, auxotóniás kontrakció alatt a feszülés csökkenhet is!). Nézzünk meg néhány példát az izotóniás kontrakcióra. A legegyszerűbb talán, ha elképzeljük, hogy fekvenyomás gyakorlatban a kinyomás (koncentrikus) fázis elején a súlyt könnyebb emelni, majd a közepén nehezebbé válik, a végén pedig ismét könnyebbnek érezzük, holott a teher tömege nem változott. A súlyzórúd mozgatási sebessége is ennek megfelelően változik (gyorsabb, lassabb, majd ismét gyorsabb). A 3.8. ábrán látható sportoló a medicinlabdát két kézzel mélytartásból, nyugalmi helyzetből dobja ki előre felfelé. A labda kiindulási sebessége nulla, majd a dobás elindításával fokozatosan növekszik, tehát a labdának gyorsulása van. Szintén auxotóniás kontrakciót végeznek az izmok, amikor guggolásból teljes erejű felugrást végzünk, vagyis saját testtömegünket nyugalmi helyzetből felgyorsítjuk. Bármilyen dinamikus mozgást is végzünk, az ellenállás legyőzéséhez különböző ízületi szögpozíciókban más és más erőt kell az izomrostoknak kifejteniük a mozgás végrehajtásához. Az egyes kontrakciótípusokat az izom feszülésének, valamint rövidülés/nyúlási sebességének tekintetében a 3.9. ábra foglalja össze.

3.8. ábra. Medicinlabda dobás két kézzel előre, nyugalmi helyzetből indítva (A: kiinduló helyzet, B: véghelyzet). A mozgásban résztvevő valamennyi izom/izomcsoport auxotóniás kontrakciót végez.
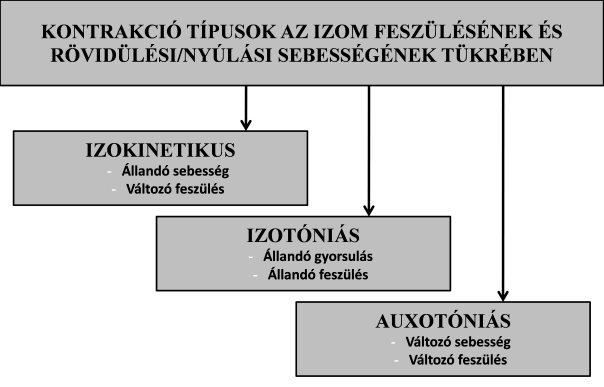
3.9. ábra. Kontrakció típusok az izom feszülésének, valamint rövidülési/nyúlási sebességének függvényében.
3.3.3. Kontrakció típusok összekapcsolása, kombinációi
Az emberi mozgások nagy része az erőkifejtési módozatokat kombinációjából áll, és a koncentrikus erőkifejtés excentrikus erőkifejtéssel párosulhat. Ha egy erősítő hatású gyakorlatot a kiinduló helyzettől a véghelyzetig elvégzünk többször egymás után, excentrikus és koncentrikus kontrakciók ciklikus ismétlődésével találkozunk (excentrikus-koncentrikus, koncentrikus-excentrikus kontrakció). A fekvőtámaszban végzett karhajlítás és nyújtás során például excentrikus erőkifejtéssel indul a gyakorlat: a vállöv izmai nyúlnak, miközben lassan lefékezik a talaj felé közelítő testet. A legmélyebb ponton a test megáll, majd a vállöv izmainak legyőző (koncentrikus) erőkifejtése következtében elindul felfelé a kiinduló helyzet felé (3.10. ábra). A két mozgásfázis közötti átkapcsolást excentrikus-koncentrikus átmenetnek (eccentric-concentric transition) nevezzük. Laboratóriumi vizsgálatokkal egyébként kimutatható, hogy az excentrikus-koncentrikus átmenet nem egy pillanat alatt történik, hanem az izomban van egy nagyon rövid ideig tartó, milliszekundum nagyságrenddel mérhető izometriás feszülés is. Természetesen az egyes kontrakció típusok kapcsolása fordított sorrendben is előfordulhat. Például guggoló támaszból emelkedés állásba (koncentrikus fázis) és ereszkedés vissza guggoló támaszba (excentrikus fázis).
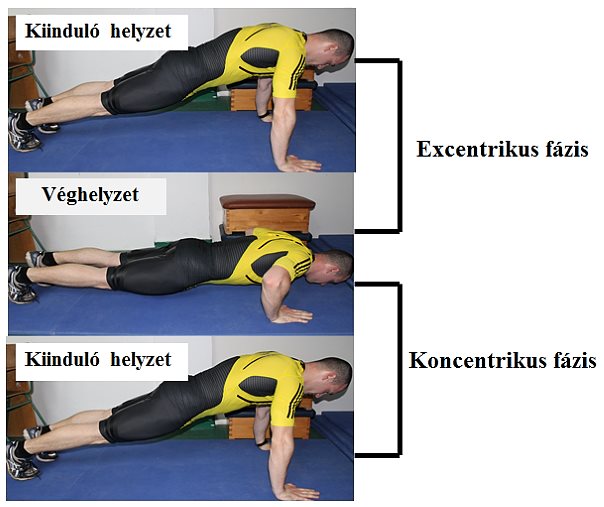
3.10. ábra. Kontrakció típusok kapcsolása fekvőtámaszban végrehajtott karhajlítás és nyújtás alatt (karhajlítás = excentrikus kontrakció fázisa, karnyújtás = koncentrikus kontrakció fázisa).
Vannak olyan természetes mozgásformák, ahol az excentrikus és koncentrikus erőkifejtések gyorsan és ciklikusan váltják egymást. Ilyen például a futómozgás, vagy a sorozatugrás (pl. szökdelések, ugrókötél gyakorlatok). Mindkét mozgásnál talajfogásnál excentrikus, az elrugaszkodásnál koncentrikus kontrakciót végeznek az izmok (3.11. ábra). Azt a kontrakció típust, ahol az excentrikus és koncentrikus fázisok nagy sebességgel követik egymást, a biomechanika tudományban nyújtásos-rövidüléses kontrakciónak nevezik. A nyújtásos-rövidüléses kontrakciónak két nagyon fontos izommechanikai jellemzője van: 1. az izom hirtelen megnyújtásakor, vagyis az excentrikus fázisban az izom elaszticitásának következtében elasztikus energia tárolódik, amely a koncentrikus fázisban hasznosul, 2. másrészt, ha az excentrikus fázis alatti nyújtás nagy sebességgel történik, akkor az izomban kiváltódik a nyújtási (miotatikus) reflex, amely szintén segíti a koncentrikus fázis munkavégzését. A kutatók már évtizedekkel ezelőtt megállapították, hogy a fenti mechanizmusok miatt az izom a koncentrikus fázisban nagyobb erő kifejtésére, illetve nagyobb mechanikai munkavégzésre képes akkor, ha azt viszonylag nagy sebességű excentrikus fázis előzi meg (Cavagna és mtsai 1968) Mindkét mechanizmust részletesebben később tárgyaljuk.
A nyújtásos-rövidüléses kontrakció, mint ahogyan már említettük a fentiekben, főleg a sorozatugrásokra jellemző. Az olyan gyakorlatokat, amelyek ilyen reaktív ugrásokat tartalmaznak, plyometriás gyakorlatoknak nevezzük. A plyometria görög eredetű szó jelentése (plyo = nagyobb, több; meter = távolság) is arra utal, hogy egy előzetes excentrikus előfeszítésből nagyobbat tudunk ugrani, mint előfeszítés nélkül.

3.11. ábra. A mélybeugrás-felugrás a plyometriás edzésmódszer egyik közkedvelt gyakorlata. Az ugrás során az izmok nyújtásos-rövidüléses kontrakciót végeznek. A = kiindulóhelyzetből, B = excentrikus-koncentrikus átmeneti pillanat, C = felugrás. Megjegyzés: az excentrikus (súlypont süllyesztése) és koncentrikus (súlypont emelése) fázisok gyorsan zajlanak le.
Végül meg kell említenünk, hogy vannak olyan mozgások is, amelyeknél statikus és dinamikus kontrakciók kapcsolódnak. Például amikor a sprintfutó atléta térdelőrajtot hajt végre, a „vigyázz” helyzet felvételekor (csípő emelése és megtartása mozdulatlanul) az alsó végtag izmai izometriásan előfeszülnek, majd a rajt indításakor koncentrikus kontrakciót végeznek az elrugaszkodáshoz. Úszók rajtkőröl végzett rajtolásakor hasonlóképpen izometriás és koncentrikus kontrakciók összekapcsolása történik. Ellenben, amikor például a tornász leugrást végez valamelyik szerről és hajlított állásba érkezik, excentrikus és izometriás kontrakciók összekapcsolásával találkozunk.
3.3.4. Az excentrikus és koncentrikus kontrakció sajátosságai
Koncentrikus és excentrikus kontrakciók önmagukban ritkán figyelhetők meg az emberi mozgásoknál. Egyes sportágakra jellemző a „tiszta” koncentrikus erőkifejtés, mint például a kerékpározásnál, az úszásnál, sífutásnál, evezésnél, vagy akár a vízilabdánál. Ezeknél a sportoknál ugyanis nincs „fékező”, csak „legyőző” izomműködés. Az emelkedőn, vagy lépcsőn felfelé gyaloglásnál is inkább a koncentrikus erőkifejtés dominál. Ezzel szemben, ha lefelé gyalogolunk, főleg excentrikus kontrakciót végeznek izmaink. Fontos megjegyeznünk, hogy a koncentrikus kontrakció köztudottan magasabb energiaigény ű, mint az excentrikus. Ebben a fejezetben már ismertettük, hogy a valós erőkifejtés koncentrikus izomműködés alatt az aktin és miozin molekulák közötti kereszthidak kialakulásától, és a molekulák konformáció-változásától jön létre, megvalósítva az izom rövidülését. Viszont az excentrikus izomműködés alatt, mikor az izomrostok valamilyen külső erő hatására megnyúlnak, az aktomiozin kötések inkább mechanikai hatásra bomlanak szét, minthogy végigmenjenek a normál ATP-függő szétkapcsolódáson (Flitney és Hirst 1978). Ezt az elméletet alátámasztja az a megállapítás, hogy az excentrikus kontrakcióban igen gyakran kisebb elektromos ingerlésre van szüksége az izomnak ugyanakkora erőkifejtéshez, mint a koncentrikusban (Grabiner és mtsai 1995). Vagyis a fentiekben említett sportágaknál és mozgásformáknál, ahol a koncentrikus izomműködés dominál, jelentős ATP igénnyel kell számolnunk, tehát a tevékenység végzése erős fáradás okozhat. A sportág űzőinek pedig magas szintű aerob és anaerob állóképességre kell szert tenniük.
Az excentrikus kontrakcióra másik sajátossága, hogy a kontrakció alatt rendkívül nagy feszülés érhető el az izomban, nagyobb, mint egy izometriás, vagy koncentrikus kontrakcióban (Doss és Karpovich 1965). Az izom feszülésének megnövekedése (ami akár 1,2-1,8-szoros is lehet) két tényezőnek tudható be: 1. Az elasztikus elemek ellenállásának növekedése a megnyújtó erővel szemben; 2. Új kereszthíd kapcsolatok létrejötte, azaz az izom aktivációs szintjének növekedése, amely a nyújtásos reflex (lásd a későbbiekben). miatt következik be akkor, ha az excentrikus kontrakció nagy sebességgel megy végbe. Van még egy elképzelés, amit Edman írt le 1999-ben. A nyújtás hatására csökken a rostok vastagsága és ezért megnő az ozmotikus nyomás és a rost tónusa.
Mindez olyan sejtszintű mikrosérüléseket okozhat az izomban, mint például a sejtmembrán, a t-tubulusok, a myofibrillumok, a szarkoplazmatikus retikulumok, vagy akár a mitokondriumok sérülése (Fridén és Lieber 1992), amely a közismert „izomláz” kiváltó oka.
ÖSSZEFOGLALÁS
- Ingerlés-kontrakció alatt azt a folyamatot értjük, amely szerint a motoneuronokból kiinduló akciós potenciálok bejutnak az izomrostokbelsejébe és ott kalcium kiáramlást okozva aktin-miozin kötéseket (kereszthidakat) hoznak létre.
- Az izom nagy frekvenciával történő ingerléskor tartós összehúzódásra képes, melyet komplett tetanusznak nevezünk. A kisülési frekvencia mértéke szabályozza a kontrakció erejét.
- Az elektromechanikai késés az ingerület megjelenése és az izom mechanikai válasza között eltelt idő. A késési időt a biokémiai folyamatok időigénye mellett a soros elasztikus elemek nyúlása is befolyásolja.
- Az izom hosszváltozásának tekintetében háromféle kontrakció típust különítünk el: izometriás (az aktív izom hossza nem változik), excentrikus (az aktív izom nyúlik) és koncentrikus (az aktív izom rövidül)
- Az auxotóniás kontrakció a legtermészetesebb kontrakció. Auxotóniás kontrakció alatt az izom hosszváltozási (rövidülés, nyúlás) sebessége és az izom feszülése is állandóan változik.
- Minden kontrakció a kontraktilis és az elasztikus izomrészek kölcsönhatásának eredménye. A kontraktilis elemek az aktív erőkifejtésért felelnek. Az elasztikus elemek a passzív erőkifejtésért, vagy aktív erőkifejtés (kontrakció) közben a kontraktilis elemek által kifejtett erő közvetítéséért felelnek.
- Ha az aktív izmot erőteljes, gyors nyújtás éri, nem csak a passzív elasztikus elemek feszülése, de az izom akiv feszülése is növekszik. Amennyiben az izom nyújtását időkésés nélkül követi az izom rövidülése, az izom mechanikai munkavégzése megnövekszik a koncentikus kontrakció során.
Irodalomjegyzék
Cavagna G, Dusman B, Margaria R (1968) Positive work done by a previously stretched muscle. Journal of Applied Physiology, 24, 21-32.
Cavanagh PR, Komi PV (1979) Electromechanical delay in human skeletal muscle under concentric and eccentric contractions. European Journal of Applied Physiology Occup Physiol 42, 159–163.
Cook R (1997) Actomyosin interaction in striated muscle. Physiological Reviews, 77, 671-697.
Doss WS, Karpovich PV (1965) A comparison of concentric, eccentric, and isometric strength of elbow flexors. Journal of Applied Physiology, 20, 351-353.
Edman KAP (1999) The force bearing capacity of frog muscle fibres during stretch: its relation to sarcomere length and fibre width. Journal of Physiology, 519.2, 515-526.
Flitney FW, Hirst DG (1978) Cross-bridge detachment and sarcomere “give” during stretch of active frog’s muscle. Journal of Physiology London, 276, 449-465.
Fridén J, Lieber RL (1992) Structural and mechanical basis of exercise-induced muscle injury. Medicine and Science in Sports and Exercise, 24, 521-530.
Grabiner MD, Owings TM, George MR, Enoka RM (1995) Eccentric contractions are specified a priori by the CNS. Proc. Congr. Int. Soc. Biomech. Jyvaskyla, Finland July 2–6, 338–339.
Huxley AF (1974) Muscle contraction (Review Lecture). Journal of Physiology, 243, 1-43.
Mines GR (1913) On functional analysis of the action of electrolytes. Journal of Physiology, 46, 188-235.
Huxley AF, Simmons R (1971) Proposed mechanism of force generation in striated muscle. Nature, 233, 533-538.
Klavora P (2007) Foundations of kinesiology: Studying human movement and health. Sport Book Publisher, Toronto, New York.
Lymn RW, Taylor EW (1971) Mechanism of adenosine triphosphate hydrolysis by actomyiosin. Biochemistry, 10, 4617-4624.
Peachey LD (1965) The sarcoplasmic reticulum and transverse tubules of the frog’s sartorius. Journal of Cell Biology, 25, 209-231.
Ringer S (1883) A further contribution regarding the influence of different constituents of the blood on the contraction of the heart. Journal of Physiology, 4, 29-42.
Stein LA, Shwartz Jr RP, Chock PB, Eisenberg E (1979) Mechanism of the actomyosin adenosine triphosphatase. Evidence that adenosine 5'-triphosphate hydrolysis can occur without dissociation of the actomyosin complex. Biochemistry, 18, 3895–3909.
| « Előző fejezet | Tartalomjegyzék | Következő fejezet » |
Események
Jelenleg nincs aktuális esemény.