TÁMOP-4.1.2.E-13/1/KONV-2013-0012
"Sporttudományi képzés fejlesztése
a Dunántúlon"
- -> Tananyagfejlesztés
- -> Mozgásszabályozás
- -> 5.












5.
5. Mozgásszabályozásban szerepet játszó legfontosabb érzékszervek működései
- 5.1. Látás
- 5.1.1. A szem felépítése
- 5.1.2. A szem optikai rendszere/A szem fénytörő közegei
- 5.1.3. A látás perifériás mechanizmusa/A retina felépítése
- 5.1.4. A látás perifériás mechanizmusa/A fototranszdukció
- 5.1.5. A látás perifériás mechanizmusa/A transzmitterfelszabadulás
- 5.1.6. A látás perifériás mechanizmusa/ Az ingerületterjedés iránya
- 5.1.7. A látás centrális mechanizmusa
- 5.2. Hallás
- 5.2.1. Akusztikai alapfogalmak
- 5.2.2. A hallás perifériás mechanizmusai/A külső fül szerepe
- 5.2.3. A hallás perifériás mechanizmusai/A középfül szerepe
- 5.2.4. A hallás perifériás mechanizmusai/A belső fül felépítése
- 5.2.5. A hallás perifériás mechanizmusai/A belső fül működése
- 5.2.6. A hallás centrális mechanizmusai
- 5.3. Helyzetérzékelés
A látás, a hallás érzékszervi működései, fontos szerepet játszanak a mozgásszabályozásban. A helyzetérzékelés a fej- és a szemek térbeli helyzetének reflexes szabályozásához, illetve a testtartás és az izomtónus (lásd 6.1. fejezet) kialakításához feltétlenül szükséges.
5.1. Látás
Az állatvilág és természetesen maga az ember is a külvilágról szóló, a külvilágban történt eseményeket a látható fény feldolgozása alapján szerzi meg. A fényinger ingerületté alakítására szolgáló érzékszerv a szem. A szem a felvett jeleket továbbítja a központi feldolgozó apparátusba, az agyba.
5.1.1. A szem felépítése
A szem szerkezeti felépítésnek (5-1. ábra) központjában a gömbölyű szemgolyó áll, melynek falát három réteg burkolja, ezek kívülről-befelé haladva a következők:
-
külső réteg, aminek hátsó része a fehér színű ínhártya (sclera), míg elülső része az átlátszó
szaruhártya (cornea). Az ínhártya felszínéhez tapadnak a szemmozgató izmok. Az embernél mindkét szemgolyót különálló izmok mozgatják,
melyek a tér mindhárom irányába képesek elmozdítani,
*Kiegészítés: Az ínhártya fehér színe a szaruhártya körül a kötőhártyán is áttűnik. Ez adja a „szem fehérét”. -
középső réteg,
mely erekkel gazdagon átszőtt, ezzel biztosítva a szem belső nedvkeringését és a belső réteg oxigénellátását, ill. táplálását, három részből áll:
-
hátsó 2/3-át a sötét színű érhártya (choroidea) borítja,
*Kiegészítés: Egyes állatoknál (pl.: macska), az érhártya egy fényvisszaverő réteget (tapetum lucidum) tartalmaz. Ebben a rétegben található sejtek guanint tartalmaznak, mely a beeső fényt sötétben zöldesen fluoreszkálva veri vissza. - az érhártya előrefelé a sugártestben folytatódik, ami a csarnokvizet termeli. A csarnokvíz víztiszta, az agy-gerincvelői folyadékhoz hasonló, mely a szemcsarnokokat tölti ki. A csarnokvíz elvezetése a csarnokzugon keresztül történik. Ha az elvezetés nem megfelelő, a szemben uralkodó nyomás fokozódik. A sugártest simaizom elemeket is tartalmaz, melyek lencsefüggesztő rostokon keresztül kapcsolódnak a rugalmas szemlencséhez. A lencsefüggesztő rostok a szemlencse mozgatásában játszanak szerepet,
- az érhártya elülső része a szivárványhártya (iris), melynek középső kerek része a 4mm átmérőjű pupilla, mely erős fény hatására összeszűkül, sötétben kitágul, így szabályozva a beengedett fény mennyiségét.
-
hátsó 2/3-át a sötét színű érhártya (choroidea) borítja,
- belső réteg, a tényleges ingerfelvevő réteg, az ideghártya (retina), mely fejlődését tekintve ugyanabból a szövetből (ektoderma) származik, mint amiből az agy fejlődik.
A retinában egyrészt fényérzékelő receptorok (csapok és pálcikák), valamint ingerületet elvezető neuronok találhatók (lásd 5.1.6. fejezet). A retinát egyrészt belülről az üvegtest, kívülről az érhártya határolja. A retinának azt a részét, ahol a látóideg a szemből kilép, vakfoltnak nevezzük. A neve onnan adódik, hogy ez a hely csapokat és pálcikákat nem tartalmaz, nincs fényérzékelés. Ettől oldalra (embernél körülbelül 4-5mm) található az éleslátás helye, a sárgafolt. A sárgafolt szerkezete eltér a retina általános szerkezetétől, mert ezen a részen kizárólag csapok találhatók.
A szemgolyó nagy részét az üvegtest alkotja. Az üvegtest előtt helyezkedik el a korábban már említett szemlencse, mely a szembe jutó fénysugarak szabályozásában játszik kiemelkedő szerepet.

5-1. ábra: A szem felépítésének vázlata
5.1.2. A szem optikai rendszere/A szem fénytörő közegei
A látás elengedhetetlen feltétele, hogy a külvilágról szóló információ megjelenjen a szem ideghártyája, a retina fényérzékelő elemein. A szem mindig a külvilág kicsinyített, fordított állású, valódi képét vetíti a retinára.
A szembe érkező fénysugárnak a retináig négy közegen kell keresztülhatolnia (szaruhártya, csarnokvíz, lencse, üvegtest). Először a szaruhártyán halad keresztül, ami egyúttal az első törőközeg. A másik nagy fénytörő rész, a pupilla mögött elhelyezkedő lencse. Az emberi szem fénytörésében a legnagyobb szerepet a szaruhártya játssza. A beeső fénysugarakat a törőközegek úgy törik meg, hogy a sugarak a retina sárgafoltján egyesüljenek (5-2. ábra/felső kép).
Távolra nézéskor a lencsefüggesztő rostok feszesek, a sugártest izmai elernyedtek, a lencsét viszonylag lapos állapotban tartják. Közelre nézéskor a sugártest izmai összehúzódnak, a lencsefüggesztő rostok feszülése csökken, a lencse rugalmasságának köszönhetően domborúbb lesz. Ezt a folyamatot, amikor a szem alkalmazkodik a különböző távolságokhoz, akkomodációnak nevezzük. Nagyságát dioptriában (D) adhatjuk meg.
Már fiatalkorban is előfordulhat, hogy a beérkező fénysugarak a szaruhártya megváltozott törőképessége miatt, a retina előtt (rövidlátás, myopia), ill. a retina mögött (távollátás, hypermetropia) egyesülnek (5-2. ábra/középső és alsó kép). Az előbbi szórólencsével, az utóbbi gyűjtőlencsével korrigálható.

5-2. ábra: A normál-, a rövid-, ill. a távollátás sematikus ábrája
*Kiegészítés: A szem optikai rendszere csak abban az esetben működik kifogástalanul, ha a fő törőközegek (szaruhártya és lencse) egymástól és retinától való távolságuk állandó. Az állandó értéken tartást a szem belnyomása biztosítja. A nyomást a csarnokvíz folyamatos termelődése és felszívódása tartja állandó értéken. A termelődés és felszívódás egyensúlyának megbomlása zöldhályoghoz vezethet, ami a retina károsodásával jár. A zöldhályog elnevezés helytelen, hiszen hályog nem képződik, és a szem nem látszik zöldnek.
5.1.3. A látás perifériás mechanizmusa/A retina felépítése
A nyugvó szem esetén a kép a retinára vetül. Embereknél „inverz” szemekről beszélhetünk, mert a fényfelfogó része a fény irányával megegyezően helyezkedik el, és csak a visszavert fénysugarakat képes felfogni. A retina réteges szerkezetű (10 réteg), melynek egyik része a pálcikák és csapok rétege. A két fotoreceptorsejt eltérő arányban (körülbelül 120 millió pálcika, 10 millió csap) van jelen (5-3. ábra). A hosszabb, henger alakú külső szegmenssel rendelkező pálcikasejtek, nagy fényérzékenységűek. Már gyenge fény mellett is működnek. Ők felelősek a szürkületi, az ún. „szkotópiás” látásért. A nappali látás, a „fotópiás” látás a rövidebb, kúp alakú csapok működéséhez kötődik. A csapok fényérzékenysége kisebb a pálcikákénál, mégis az ő működésük teszi lehetővé a színek érzékelését. Mindkét fényérzékelő sejt különböző feladatot ellátó részre tagolódik (5-3. ábra). Külső szegmensükben korong alakú membránrendszer figyelhető meg, melyek fényérzékeny fehérjemolekulákat, az ún. fotopigmenteket tartalmazzák. A belső szegmentumban találjuk a sejtmagot, ill. a mitokondriumot, mely a sejt anyagcsere-folyamatához elengedhetetlen. A belső szegmens a szinaptikus végződésbe megy át, itt helyezkednek el az ingerületátvivő anyagot (transzmitter) tartalmazó hólyagocskák.
5.1.4. A látás perifériás mechanizmusa/A fototranszdukció
Fény hatására a fotoreceptorsejtekben fotokémiai folyamat indul meg. A pálcikák fotopigmentje a rodopszin (látóbíbor). A rodopszinban erős kémiai kötéssel (kovalens kötés) kapcsolódik a retinal, ami az A-vitamin származéka. A retinalmentes fehérjerészt opszinnak nevezzük. Ha fény éri a fotoreceptorsejtet, a retinal szerkezete megváltozik. A változás különbőző folyamatokon keresztül egy nukleotid, a ciklikus guanozin-monofoszfát (cGMP) szintjét csökkenti. A cGMP szint csökkenése a látás alapvető lépése.
*Kiegészítés: 1.) A kémiai reakciót követően az átalakult retinal leválik a fehérjéről és elhagyja a sejtet. A sejtből kijutott retinalt a szomszédos sejtek felveszik és A-vitaminná, majd eredeti rodopszinná alakítják. A szervezet nem képes A-vitamint előállítani. Ennek hiányában a sötétben látás képessége szűnik meg, amit „farkasvakságnak” nevezünk. 2.) A rodopszin visszaalakítása sötétben lassú, percekben mérhető folyamat. Ez az oka, hogy ha fényről a sötétbe megyünk, a látásunk (pálcikalátás) csak percek múlva áll vissza.

5-3. ábra: A fényérzékelő receptorok felépítése
5.1.5. A látás perifériás mechanizmusa/A transzmitterfelszabadulás
A fotoreceptorsejtek nyugalmi membránpotenciálja sötétben pozitívabb (-40mV, 5-4. ábra), mint más idegsejtek membránpotenciálja (-70-90mV). Ez az emelkedett membránpotenciál azt jelenti, hogy sötétben a fotoreceptorsejtek depolarizált állapotban vannak. Ennek oka, hogy sötétben a sejt külső szegmentumában a magas cGMP-szint hatására nyitottak a Na+-csatornák. A Na+ a nagyobb koncentrációjú helyről a kisebb felé, azaz a sejtbe áramlik, ezáltal depolarizációt okoz. A depolarizáció hatására a szinaptikus végződésből neurotranszmitter anyag, jelen esetben glutamát szabadul fel. Fény hatására a cGMP-szint lecsökken, a Na+-csatornák bezárulnak. Eközben a folyamatosan áramlik ki a sejtből a K+, a sejt hiperpolarizálódik, a potenciálkülönbség értéke eléri a -70 mV-ot. A hiperpolarizáció következtében a szinaptikus végződésből a glutamát leadása csökken.

5-4. ábra: A fotoreceptor sejtek transzmitter ürítésének vázlata
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 935. oldal, 37-31. ábra alapján/
5.1.6. A látás perifériás mechanizmusa/ Az ingerületterjedés iránya
A retinában a fotoreceptorsejtek mellett különböző idegsejteket lehet elkülöníteni: bipoláris (kétnyúlványú) , horizontális, amakrin, ill. dúcsejteket (5-5. ábra). A fotoreceptorsejtekről a potenciálváltozást a bipoláris idegsejtek vezetik tovább a retinában. A bipoláris idegsejtek nyúlványai a dúcsejtekhez kapcsolódnak. Ezeknek a dúcsejteknek az axonjai alkotják a látóideget (nervus opticus).

5-5. ábra: Az ingerületterjedés útja
5.1.7. A látás centrális mechanizmusa
A látóideg a vakfolton keresztül, a szemgolyó hátsó részén lép ki. A látóideg egy ideig változatlanul halad, majd eléri a középagyat (chiasma opticum), ahol egy része átkereszteződik (lásd 6-1. ábra). A szemidegek kereszteződése után márlátópályáról beszélhetünk, ami a talamusz CGL (corpus geniculatum laterale) magjába viszi az ingerületet. Innen átkapcsolódás után az információ az elsődleges látókéregbe jut (Brodmann 17), ami a nyakszirtlebenyben található (5-6. ábra). Az elsődleges látókéregből a vizuális információ az agykéreg egyéb területeire (másodlagos, ill. harmadlagos látókéreg) is eljut. A látásélmény a kéregrészek együttműködésének eredménye.

5-6. ábra: A vizuális információk iránya a nagyagykéregben
5.2. Hallás
5.2.1. Akusztikai alapfogalmak
A hang a levegő longitudinális rezgése, azaz egymást meghatározott frekvenciával követő légsűrűsödések és légritkulások sorozata. A hangokat hangmagassággal és hangerővel szokás jellemezni. A hangmagasság a rezgés ciklusainak periódusidejével (hullámhossz) vagy a periódusidő reciprokával, azaz a frekvenciával jellemezhető, melyet Hertz-ben (Hz) szokás megadni. Az ún. tiszta hangok egyetlen jellemző frekvenciával írhatók le, függvényen ábrázolva szinuszgörbét adnak.
A valóságban előforduló összetett hangok több, ún. harmonikussal jellemezhetők, melyek az eredeti frekvencia egész számú többszörösei és „rátevődnek” az eredeti szinuszhullámra. Így jönnek létre az eltérő hangszínek, pl. két különböző hangszer is így szólalhat meg másféle hangon annak ellenére, hogy éppen ugyanazt a zenei hangot szólaltatják meg. Az emberi fül a 20-20000Hz frekvenciájú hangokat képes észlelni, a 20Hz alatti frekvenciájú hangot infrahangoknak, míg a 20000Hz felettieket ultrahangoknak nevezzük. Hallószervünk az 1000-4000Hz-es tartományban a legérzékenyebb. Érdekes módon ennél kicsit alacsonyabb tartományba esnek a beszédhangok (80-1000Hz). Feltételezhetően azért, mert az evolúció során a hallás nem elsősorban a beszéd megértésére fejlődött ki.
A hang erősségét (hangintenzitás) a levegő rezgése miatt létrejövő nyomásváltozás nagyságával jellemezhetjük. Ez tulajdonképpen a hangot leíró szinuszfüggvény amplitúdójának nagysága. A hangerőt leggyakrabban mégsem a nyomás leírására szolgáló Pascal (Pa) fizikai mértékegységgel, hanem decibelben (dB) mérjük. A legfontosabb különbség a kettő között az, hogy a decibel skálán a 0dB pontban van mérhető hangnyomás. Ez 2000Hz-es frekvencián éppen a hallásküszöb értéke, tehát az a hangerő, amit egy egészséges ember még éppen meghall. Értéke:
0db = 2x10-5Pa = 20mPa.
Ez azonban csak 2000Hz-nél érvényes, ugyanis alacsonyabb és magasabb frekvenciatartományokban fülünk kevésbé érzékeny, így a hallásküszöb értéke is egyre magasabb dB-tartományokat vesz fel (5-7. ábra).

5-7. ábra: Az emberi fül hallástartományának hangmagasság (frekvencia) és hangerő (hangnyomás) függése. Pirossal jelezve a legérzékenyebb tartomány
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 913. oldal, 37-10. ábra alapján/
Fülünk a 40-80dB tartományokban hall a legjobban, az ennél magasabb hangintenzitás értékek már kellemetlenek, 120dB felett pedig már nem hangot, hanem fájdalmat érzünk.
5.2.2. A hallás perifériás mechanizmusai/A külső fül szerepe
A fülkagyló és a külső hallójárat alkotják a külső fület. Rezonátorként működve a levegőrezgést a dobhártyáig vezetik. A fülkagyló hangerősítő szerepe elhanyagolható, a hangirány meghatározásában játszhat inkább szerepet.
5.2.3. A hallás perifériás mechanizmusai/A középfül szerepe
A középfül a dobhártyától a belső fül ovális ablakáig vezeti a rezgéseket. Szerepe az, hogy megfelelő erősítést adjon ahhoz, hogy az eddig a kisebb sűrűségű levegőben könnyebben terjedő rezgés a belső fülben, mint folyadékrezgés folytathassa útját. Ezt a folyamatot impedanciaillesztésnek nevezzük, melyet a hallócsontocskák (5-8. ábra) végzik.
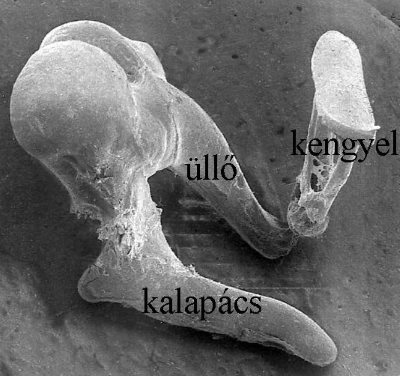
5-8. ábra: A hallócsontocskák elektronmikroszkópos képe
Megfigyelhető a kalapács „nyele” mely normálisan a dobhártyához rögzül, ill. a kengyel hosszúkás talpa mely pontosan illeszkedik a csiga ovális ablakához.
Az impedanciaillesztés során a levegőrezgés mintegy 22-szeres erősítést kap a kalapács-üllő-kengyel által kifejtett emelőhatásnak köszönhetően. Az így felerősített jel már képes a belső fülben érzékelhető folyadékrezgésként tovaterjedni az ovális ablakból kiindulva.
*Kiegészítésként elmondható, hogy a hangok nem csak a dobhártya közvetítésével, hanem a koponyacsontok rezgése által is eljuthatnak a belső fülbe. Mivel beszéd közben a koponyacsontok rezonátorként működnek, saját beszédhangjaink két úton juthatnak el a csigába, így lehet az, hogy másnak halljuk saját beszédhangjainkat, mint egy külső személy. Ez az oka annak is, hogy gyakran idegennek tűnik, pl. a rögzített beszédhangunk visszahallgatása.
5.2.4. A hallás perifériás mechanizmusai/A belső fül felépítése
A belső fülben található a csiga (cochlea). A csiga belső ürege három csatornára oszlik, melyek egymás felett „emeletszerűen” helyezkednek el. A folyadékrezgések az ovális ablaktól a scala vestibuli-ban terjednek a csiga csúcsa felé. Itt a scala vestibuli megszakítás nélkül folytatódik a scala tympani-ban, mely visszavezet a csiga alapjához és a kerek ablaknál ér véget (5-9. ábra és 5-10. ábra).
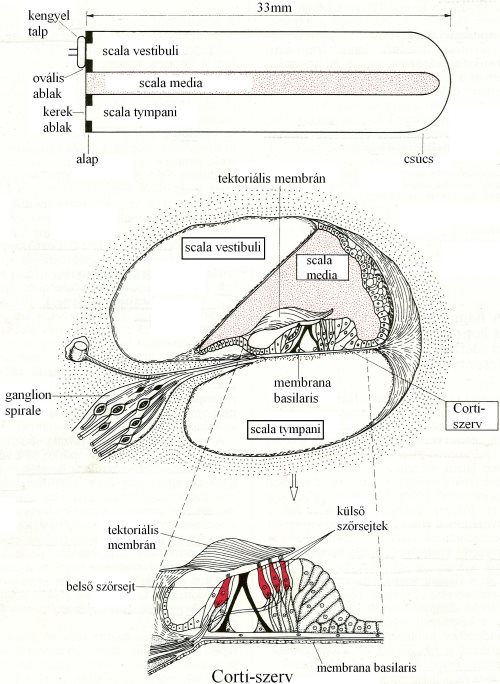
5-9. ábra: A csiga három belső üregének elhelyezkedése és a Corti-szerv felépítése
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 916. oldal, 37-13. ábra alapján/

5-10. ábra: A csiga elektronmikroszkópos képe
A scala vestibuli és a scala tympani közrefogja a csiga középső üregét a scala mediát, mely a hallás receptorzónáját a Corti-szervet tartalmazza (5-9. ábra). A Corti-szerv a scala tympanit és a scala mediát elválasztómembrana basilarison fekszik. Ezen találhatóak a három sorban fekvő külső-, és az egy sort alkotó belsőszőrsejtek. A szőrsejtek közötti teret támasztósejtek töltik ki. A külső szőrsejtek a tetejükön lévő csillókkal (cilliumok) a föléjük hajló tektoriál is membránhoz vannak rögzülve, míg a belső szőrsejtek csillói a scala media folyadékába nyúlnak. Mind a külső-, mind a belső szőrsejtek alapjuknál szinapszist képeznek a ganglion spirale bipoláris neuronjaival, melyek a hallóideget alkotva a szőrsejtekben keletkezett ingerületeket továbbítják az agytörzs felé (részletesen ld. lejjebb).
5.2.5. A hallás perifériás mechanizmusai/A belső fül működése
A csiga a benne terjedő folyadékrezgéseket frekvencia (hangmagasság) és intenzitás (hangerő) szerint analizálja. A folyadékrezgéseket a Corti-szervben lévő szőrsejtek elektromos jelekké alakítják (mechanoelektromos transzdukció). Az így létrejött akciós potenciálok végül a hallóideg és a hallópályák közvetítésével jutnak el a hallókéregbe, ahol a hallásérzet kialakul (5-11. ábra).
A hangok által keltett folyadékrezgések a membrana basilarison ún. vándorlóhullámokat hoznak létre. A membrana basilaris a csiga alapjánál keskeny és feszes, míg a csiga csúcsánál széles és laza. Ennek köszönhetően a magas hangok a csiga alapjánál térítik ki legjobban a membránt, míg a mély hangok a csiga csúcsánál
A nyugalmi helyzetéből kimozduló membrana basilaris „magával húzza” a tektoriális membránhoz csillóikkal kapcsoló külső szőrsejteket. A külső szőrsejteken e mechanikus inger (a csillók kitérése) hatására K+-csatornák nyílnak és K+ áramlik be a sejtbe. A beáramló K+-kationok depolarizálják a külső szőrsejteket (mechanoelektromos transzdukció, 5-11. ábra). A depolarizáció hatására átalakul a külső szőrsejtek belső szerkezete, csillóik elmozdulnak és magukkal „húzzák” a tektoriális membránt (elektromechanikus transzdukció). A külső szőrsejtekben tehát először mechanikus ingerek alakulnak elektromossá majd fordítva: a kialakult elektromos változás (depolarizáció) mechanikai folyamatot indukál. Ezt a kettős hatást nevezzük aktív bidirekcionális transzdukciós mechanizmusnak. A külső szőrsejtek hatására elmozduló tektoriális membrán már a belső szőrsejteket is ingerületbe hozza, melyek hasonlóan mechanoelektromos transzdukcióval válaszolnak (5-11. ábra). A külső szőrsejtek tehát „felerősítik” a membrana basilarison terjedő vándorlóhullám hatását, így lehetővé teszik a belső szőrsejtek számára a hanghullámok csúcsainak pontosabb detektálását. A frekvenciaanalízis során ekképpen magas hangot hallunk akkor, ha a csiga alapjánál lévő belső szőrsejtek jönnek ingerületbe, míg mélyet, ha a csiga csúcsán elhelyezkedők.
Az ingerületbe jövő szőrsejtek glutamát neurotranszmittert ürítenek az alattuk elhelyezkedő szinaptikus résbe (5-11. ábra), mely az afferens idegvégződéseken akciós potenciálok kialakulásához vezet. A hangerő (intenzitás) kódolása: ha erősebb a hallott hang, akkor nagyobb mértékben depolarizálnak a szőrsejtek, így több transzmittert ürítenek, mely végül az afferens idegvégződéseken kialakuló akciós potenciálok számát megnöveli.

5-11. ábra: A szőrsejtek mechanoelektromos transzdukciója
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 921. oldal, 37-19. ábra alapján/
5.2.6. A hallás centrális mechanizmusai
Az afferens idegvégződéseken kialakuló akciós potenciálokat a ganglion spirale bipoláris neuronjai a hallóideget (VIII. agyideg) alkotva vezetik tovább az agy felé. A hallóideg az agytörzsi nucleus cochlearis-ba jut. Az innen kiinduló axonok a középagyba jutnak, ahonnan több átkapcsolás után az azonos oldali vagy az ellenoldali talamusz CGM (corpus geniculatum mediale) magjába jut az ingerület. Végül a CGM-ből a primer hallókéregbe (Brodmann 42) kerül az információ, mely a halántéklebenyben található. A csiga, a nucleus cochlearis, és a hallókéreg bizonyos részei tonotóp vetülésűek, azaz a különböző magasságú hangoknak meghatározott helyük van. Az érzékszervek közül a hallás pályái a legösszetettebbek, többek közt a hangirány meghatározásának bonyolultsága miatt.
5.3. Helyzetérzékelés
5.3.1. A helyzetérzékelés szerve
A belső fülben a csiga mellett helyezkednek el a tér mindhárom irányába egy-egy ívet rajzoló félkörös ívjáratok (5-12. ábra). Mindegyik ívjárat az utriculus-ból indul ki és oda is tér vissza. Kiszélesedő részüket ampullának nevezzük, itt találhatóak azok a receptormezők, melyek a fej szöggyorsulását (elfordulását) detektálják. Az utriculus-ban és a hozzá kapcsolódó sacculus-ban helyezkedik el az otholitszerv, mely a lineáris gyorsulást és a fej térbeli helyzetét érzékeli.
5.3.2. A fej szöggyorsulásának detektálása
A fej elfordulásának hatására a félkörös ívjáratokban lévő belső folyadék, tehetetlensége miatt az ellenkező irányba mozdul el. Az elmozduló folyadék az ampullában található kocsonyás szerkezetű cupulát eltorzítja. A cupulába nyúlnak a szőrsejtek csillói, melyek a folyadék elmozdulásával egyező irányba kitérnek (5-13. ábra). A csillók kitérése mechanoelektromos transzdukciót okoz, hasonlóan a Corti-szervnél leírtakhoz. Az afferens idegen akciós potenciálok alakulnak ki, és végül az információ a vesztibuláris ideg közvetítésével az agytörzsi vesztibuláris magokhoz jut.

5-12. ábra: A belső fül a csigával és a félkörös ívjáratokkal
1. utriculus, 2. félkörös ívjáratok, 3. sacculus, 4. ampulla, 5. csiga, 6. vesztibuláris ideg, 7. hallóideg

5-13. ábra: A fej elfordulásának érzékelése
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 853. oldal, 35-18. ábra alapján/
5.3.3. A lineáris gyorsulás- és a fej térbeli helyzetének detektálása
Az utriculusban és a sacculusban elhelyezkedő otholitszerv képes érzékelni a lineáris gyorsulást, pl. liftben érezzük a lift gyorsulását vagy lassulását. Illetve érzékeli a fej térbeli helyzetét, pl. csukott szemmel is képesek vagyunk meghatározni a le és a fel irányokat. Az otholitszervben hasonló működésű szőrsejtek találhatók, mint az ampullákban, azzal a különbséggel, hogy csillóik nem cupulába, hanem egy hasonlóan kocsonyás szerkezetű otholitmembránba nyúlnak (5-14. ábra). Az otholitmembrán erőhatásra bekövetkező torzulását a rajta fekvő otholitkristályok (CaCO3) tehetetlensége vagy súlya okozza.

5-14. ábra: Az otholitszerv
/Forrás: Fonyó, Az orvosi élettan tankönyve, 1999., 854. oldal, 35-19. ábra alapján/
5.3.4. Hová jut a helyzetérzékeléssel kapcsolatos információ?
Mind a három ampullából, mind az otholitszervekből érkező információ a vesztibuláris idegen keresztül hagyja el a belső fület. A helyzetérzékeléssel kapcsolatos információk az agytörzsben elhelyezkedő vesztibuláris magokhoz, ill. közvetlenül a kisagyba futnak (lásd 3.6. fejezet). A vesztibuláris magok sok agyterülettel kapcsolatban állnak és a helyzet- és egyensúlyérzékelés mellett a testtartás kialakításában is részt vesznek, ezért szerepüket a következő fejezetben (lásd 6.1. fejezet) elemezzük részletesebben.
| « Előző fejezet | Tartalomjegyzék | Következő fejezet » |
Események
Jelenleg nincs aktuális esemény.